перевод на русский, синонимы, антонимы, произношение, примеры предложений, транскрипция, определение,значение, словосочетания
Варианты
(v1)Варианты
(v2)| noun | |||
| подушка | pillow, cushion, pad, chock, saddle, bearer | ||
| борт | board, bead, side, broadside, bort, cushion | ||
| диванная подушка | cushion, squab | ||
| воздушная подушка | airbag, air cushion, cushion | ||
| упругая прокладка | cushion | ||
| verb | |||
| смягчать | soften, mitigate, remit, alleviate, moderate, cushion | ||
| замалчивать | hush up, suppress, sink, cushion, suppress discussion, avoid giving publicity | ||
| подкладывать подушку | cushion | ||
| обходить молчанием | elide, pass by in silence, cushion | ||
| защищать | protect, defend, advocate, uphold, guard, cushion | ||
Предложения со словом «cushion»
I’m afraid this poor girl is a pin cushion . | Боюсь, что эта бедная девушка напоминает подушку для булавок. |
He watched her kneel awkwardly on the cushion opposite. | Он посмотрел на ее колено, неловко положенное на подушку . |
Her right hand brought out her stiletto dagger from her white obi and she placed it on the cushion in front of her. | Правой рукой она достала из — за оби узкий нож и положила его на подушку перед собой. |
He activated the air cushion and withdrew the supports, so that the crystal floated before them. | Активировал воздушную подушку и выдернул подставки, так что кристалл стал парить перед ними. |
His eloquence was arrested abruptly, as Tuppence threw a cushion at him and requested him not to talk nonsense. | Поток его красноречия был прерван запущенной Таппенс подушкой и просьбой не молоть чепухи. |
Then he placed the cushion against Barger’s head, pushed the muzzle of the gun into the fabric, and pulled the trigger. | Потом он приставил подушку к голове Баргера, утопил дуло пистолета к ткани и спустил курок. |
You need to start your life, not cushion the fall for him. | Тебе надо начать жизнь с начала, а не смягчать его удары. |
I puffed a deep breath into my cheeks, slumped back against the cushion of the booth. | Я сделала глубокий выдох и откинулась на спинку стула. |
Did you happen to find a little pin cushion in here anywhere, Father? | Вы не видели случайно подушечку для иголок, Отец? |
Mnov-korthe made no effort to cushion her fall, jumping back instead out of her way as she crashed to the floor with a grunt of pain. | Мнов — корт даже не попытался подхватить ее, наоборот, отпрыгнул подальше, когда она рухнула с болезненным стоном. |
Then I moved him to his straw cushion and went reluctantly to my afternoon lessons and chores. | Тогда я перенес его на соломенную подстилку и неохотно занялся своими дневными уроками. |
Services may act as a cushion during periods of economic recession to mitigate manufacturing swings. | В периоды экономических спадов сектор услуг может играть роль буфера, смягчая последствия резких колебаний в сфере промышленного производства. |
They should cushion families and children against deprivation which may push families to discard their children and/or children to enter exploitative trades. | Им следует позаботиться о том, чтобы семьи и дети не испытывали лишений, которые могли бы вынудить семьи отказываться от заботы за своими детьми и/или привести к эксплуатации детей. |
If possible within these constraints, place the thighs of the dummy in contact with the seat cushion . | Бедра манекена, если это возможно при данных условиях, должны соприкасаться с подушкой сиденья. |
The cavalry is on its way, armed with pinking shears and a fully loaded pin cushion . | Кавалерия направляется к тебе, вооруженная ножницами и подушками с иголками. |
Probably just stuffed the handkerchief in the cushion here as a Hail Mary bread crumb to get Voss busted. | Вероятно, просто подписанный носовой платок здесь, в подушке как хлебная крошка отчаянной попытки привести к аресту Восса. |
Yeah, I get that — a little extra cushion for the, uh. | Да, я получил, этот дополнительный валик для, Э — Э. |
Announcing the abolition of the hukou system might be easy, but it would mean little without institutions that can cushion the risks faced by rural migrant workers in China’s cities. | Объявить об отмене системы прописки «хукоу» может быть легко, но это будет играть небольшое значение в случае отсутствия институтов, которые смогут смягчить риски, с которыми сталкиваются в китайских городах рабочие — мигранты из сельской местности. |
They do not have the luxury of cobbling together fiscal-stimulus packages or rescuing ailing industries in order to cushion the shock brought about by the economic crisis. | Они не могут позволить себе роскошь направить финансовые стимулирующие пакеты или спасительные меры для промышленности, чтобы амортизировать шок, который нанес экономический кризис. |
An Australian study recently suggested jogging improves the composition of the cushion-like discs in your spine, reducing wear and tear. | Согласно недавнему австралийскому исследованию, бег улучшает строение межпозвоночных дисков, уменьшая износ и вероятность их травмирования. |
I should like to stick you full of barbed arrows like a pin cushion . | Мой милый, я хотел бы утыкать вас стрелами, как подушечку для булавок. |
The cost of imported Western goods has certainly skyrocketed for regular Russians, but those with oil incomes have more cushion against these inflationary pressures. | Конечно, для простых россиян цены на западные товары резко подскочили, однако для тех, кто получает прибыль от продажи нефти, инфляция не так ощутима. |
It funded the investment imperatives of economic development and boosted the cushion of foreign-exchange reserves that has shielded China from external shocks. | Они профинансировали инвестиционные императивы экономического развития и увеличили подушку валютных резервов, которые защитили Китай от внешних потрясений. |
Ergonomic heated seat cushion | Эргономическая подушка для сиденья с подогревом. |
Lastly, under this cushion of liquid helium lies Earth’s nucleus. | И, наконец, под оболочкой жидкого газа находится земное ядро. |
You really should have borrowed Richard Hammond’s booster cushion . | Тебе надо было одолжить детское сиденье у Хаммонда. |
I never wanted to sit on a cushion and sew a fine seam. | Я никогда не хотела сидеть на подушках и вышивать. |
He put the poker back and got the cushion out of Mother’s chair and gave it to Ben, and he hunkered down in front of the fireplace and got quiet. | Поставил, с матушкиного кресла взял подушечку , дал Бену, тот ссутулился перед камином на полу и замолчал. |
Oh, I have some news, said a pretty brunette named Leonie as she leaned over her cushion to crimp some rose petals. | Да, вы еще не знаете, — сказала хорошенькая брюнетка Леони, нагибаясь над подушечкой и гофрируя на ней розовые лепестки. |
You are afraid of life-as afraid of it as an Oriental who sits all day on a cushion smoking his hookah. | Вы боитесь жизни, боитесь, как азиат, тот самый, который по целым дням сидит на перине и курит кальян. |
This cushion was unzipped, this cover wasn’t straight, the corner of this carpet’s folded over, the books aren’t quite flat against the shelves. | Расстегнут чехол диванной подушки , покрывало лежало криво, отогнут угол этого ковра, книги на полках стоят неровно. |
Detective Sergeant Wilson deposed to the finding of the blowpipe behind the cushion of one of the seats. | Детектив — сержант Уилсон показал, что он нашел духовую трубку за подушкой одного из кресел. |
I must cushion the pull of the line with my body and at all times be ready to give line with both hands. | Я должен беспрерывно ослаблять тягу своим телом и быть готов в любую минуту опустить бечеву обеими руками. |
These three strata of the Olivine Belt repose on a cushion of helium gas condensed to a liquid state, the product of atomic disintegration. | Эти три слоя Оливинового пояса покоятся, как на подушке , на слое сгущенного, до жидкого состояния, газа гелия, получающегося как продукт атомного распада. |
I pressed my face against the cushion . | Я прижалась лицом к подушке . |
Benjamin. she said. Take that cushion away, Candace. | Бенджамин, — сказала мама. — Убери эту подушку Кэндейси. |
Take that cushion away, like I told you. Mother said. He must learn to mind. | Я сказала: убери подушку , — сказала мама. — Его надо научить слушаться. |
First there marched four officers of the jeddak’s Guard bearing a huge salver on which reposed, upon a cushion of scarlet silk, a great golden chain with a collar and padlock at each end. | Впереди шли четыре офицера гвардии джеддака и несли на пурпурной подушке большую золотую цепь с браслетами и замками по концам. |
The Archbishop of Canterbury lifted up the crown of England from its cushion and held it out over the trembling mock-King’s head. | Архиепископ Кентерберийский взял с подушки корону Англии и поднял ее над головой дрожавшего всем телом мнимого короля. |
Grandfather Smallweed immediately throws the cushion at her. | Дедушка Смоллуид незамедлительно швыряет в нее подушкой . |
Grandfather looks at the cushion but has not sufficiently recovered his late exertion. | Дедушка смотрит на подушку , но чувствует, что еще не совсем оправился после своего давешнего подвига. |
‘No, that won’t do,’ I said. ‘You’ll be content with the cushion , Master Heathcliff. | Нет, так не годится! — сказала я. — Хватит с вас подушки , мастер Хитклиф. |
She took her seat on the leather cushion and the motorcycle sped off. | Она села на кожаную подушку , и мотоцикл полетел вдоль дороги. |
Mr Goby emphasised the last point by nodding his head significantly at a cushion on the sofa. | Последние слова мистер Г оби подчеркнул многозначительным кивком в сторону диванной подушки . |
By Jove, Will, it was all I I could do to prevent myself from throwing the sofa- cushion at her. | En — богу, Уил, я едва — едва сдержался, чтобы не запустить в нее диванной подушкой ! |
I’m really not familiar enough with the cushion densities, air flow patterns and dispersion of unlight to make an informed choice. | Я действительно не слишком хорошо знаком с плотностью диванных подушек , моделью потоков воздуха и дисперсией солнечного света, чтобы сделать осознанный выбор. |
An old Pekinese sat upon a cushion on a door step, its mouth drooling. | Старый пекинес сидел на подушечке рядом с входной дверью, из полураскрытой пасти его текла слюна. |
Emily dearest, I think that cushion is a little damp. | Эмили, дорогая, боюсь, что подушечка немного влажная. |
I’m not going to buy a hemorrhoid cushion . | Я не собираюсь покупать подушечку от геморроя. |
Sit in this chair, it has a cushion . | Сядь в кресло, там подушечка . |
I wish we had watched Andy’s Magic Cushion . | l ХОТЕТЬ, чтобы мы понаблюдали Волшебная Прокладка Энди. |
I hate arriving early, and besides, we have a five or six hour cushion on the other side. | Ненавижу прибывать рано, и, кроме того, у нас есть пять или шесть часов на борту самолета. |
I was positing a theory to help cushion the blow for you. | Я придерживаюсь теории — смягчать удар. |
I’ve been using it to cushion insults for years. | Я так смягчаю оскорбления много лет. |
Miss Janis… you’re a great cushion . | Джанис, Вы прекрасно смягчаете. |
I have often thought that perhaps formal good manners may be a cushion against heartbreak. | Я часто думаю, что обрядность может смягчить муку, не дает сердцу разбиться. |
Are you bringing flowers to a patient to cushion the blow of a terminal diagnosis? | Вы несёте цветы пациенту, чтобы смягчить потрясение от смертельного диагноза? |
Well, you know, a young person has got a bit of fat in… on them, something to cushion the splat. | Ну , знаешь, у молодого человека есть немного жира, что — то, чтобы смягчить звук шлепка. |
We call it the Tideymans test, because only Tideymans carpets have the absorbency and luxurious shag pile bounce that can cushion a fall from even the highest of buildings. | Сейчас мы на ваших глазах проведем уникальный тест Тайдимена, ибо только ковры Тайдимена обладают длинным, толстым упругим ворсом, способным смягчить падение даже с верхних этажей. |
as if figuring it out will somehow cushion the blow. | Как будто предсказания могут смягчить удар. |












перевод на русский, синонимы, антонимы, произношение, примеры предложений, транскрипция, значение, словосочетания
Two brocade cushions faced each other on the tatamis. | На татами друг против друга были положены две парчовые подушки . |
The desks would be painted and the chairs would have cushions . | Парты были бы разукрашены, а на стульях были бы подушки . |
Near it there is a sofa with some cushions . | Возле него находится диван с подушками . |
He opened the rear door and I got in and sank down into the cushions and George slid under the wheel and started the big car. | Он открыл заднюю дверцу, и я забрался на мягкое сидение. |
Looks like low-grade foam rubber, the kind used in inexpensive throw pillows, chair cushions , and cheap furniture. | Похоже на низкосортный поролон, который используется в дешевых подушках , обивке в стульях и мебели. |
Then she bent over the cushions , reached out her hand, and from the couch arose the form of a young girl, fresh and beautiful as a May morning. | Потом она протянула руку и помогла подняться девочке, свежей и прекрасной, как майское утро. |
We often passed the shop, and every time George paused and examined those cushions . | Мы часто проходили мимо, и Джордж каждый раз останавливался и рассматривал их. |
On one of the low platforms lay what looked to be furry animal hides, cushions , and a low table with bowls of food and pitchers and goblets of drink. | На невысокой платформе, устланной мохнатыми звериными шкурами, лежали подушки , а рядом находился маленький столик. |
Korba was brought in, seated at a low table with cushions beside it on the chamber floor below the dais. | Корбу ввели, усадили на подушки возле низкого стола, неподалеку от подножия трона. |
Not a toothpick, not the spare change between the couch cushions , not the panties out of the hamper. | Ни зубочистку, ни мелочь между диванными подушками , ни даже трусики из корзины. |
She sank into the easy chair, groaning as the cushions swallowed up her heavily pregnant frame. | Она опустилась в кресло и застонала, когда ее тяжелое тело утонуло в мягких подушках . |
Its silken cushions and hangings had been stolen or destroyed by moth and mouse and mold. | Шелковые подушки и занавеси были украдены или сожраны молью, мышами и плесенью. |
They were puffy with edema, like little foot-shaped air cushions . | Из — за отеков ступни были похожи на маленькие пухлые подушечки . |
And a pair of bolster cushions , and a fake-fur throw. | И пару подушек , и покрывало из искусственного меха. |
An old couch stood in one corner of the kitchen, its springs flowering through the rotting cushions . | Угол кухни отгораживал старый диван, который топорщился вылезшими сквозь истлевшую обивку пружинами. |
They had a pile of cushions and pillows, which they dumped on the deck. | Они принесли гору подушек и свалили их на палубу. |
There were seats with hand-embroidered cushions on them, and there were tapestries covering the windows and the doors. | На сиденьях лежали украшенные ручной вышивкой подушки , двери и окна закрывали гобелены. |
Bedding, blankets, pots, all lay scattered across the stone floor amid slashed cushions and pillows. | Распоротые и располосованные подушки и одеяла валялись на каменном полу вперемешку с осколками разбитой посуды. |
And the house has the same smell of pot-pourri, flowers, tobacco, books, and sofa cushions . | В доме стоит все тот же смешанный запах цветов, табака, книг и диванных подушек . |
The seats had white velvet cushions upon them and the cushions were splendidly embroidered with threads of gold. | На сиденьях были бархатные, белые, расшитые золотом подушки . |
The man behind the desk had to be sitting either in a very tall chair or atop a heap of cushions , for he couldn’t have stood much over four feet. | Человечек за столом сидел на очень высоком стуле или на высокой стопке подушек . |
Well, I don’t have it, so start pulling up pillow cushions . | У меня его нет, так что начинай перетряхивать подушки . |
This versatile, cost-efficient system produces adjustable height cushions for various sized applications. | Универсальные и экономичные системы выпускают регулируемые по высоте валики требуемых размеров. |
When the Prestige sank off the coast of Spain, there were big, floating cushions the size of sofa cushions of emulsified oil, with the consistency, or the viscosity, of chewing gum. | Когда нефтяной танкер Престиж затонул у берегов Испании, по воде плавали большие подушки эмульгированной нефти с вязкостью и тягучестью жевательной резинки. |
In 2008-09, the country’s ample cash and fiscal cushions were used to cope with the crisis. | В 2008 — 2009 гг., чтобы справиться с кризисом, были использованы обширные денежные массы и бюджетные резервные запасы страны. |
Zoe sat amidst numerous cushions on a wide divan, her feet tucked up under her. | Зоя Монроз сидела, подобрав ноги, на широком диване среди множества подушечек . |
Sandwiches had a tendency to become mislaid during these pauses, and to vanish down between the cushions of the seats. | В такие минуты положенные куда попало бутерброды исчезали, проваливались между подушками сидений. |
ugly, old and surly, always glued to her TV while her fat cat dozes on cushions covered with crocheted cases in a stink of bean stew. | уродливая, старая и неприветливая, всегда прикованная к своему телевизору в то время как её толстый кот дремлет на диванных подушках с трикотажными чехлами в зловонии бобового тушеного мяса. |
frayed cushions and moldy walls. | Вытертые подушки . Стены, поросшие плесенью. |
Up until the lunch break, everybody in Finance and Accounting fidgeted on their stools and cushions , dying of curiosity. | До обеденного перерыва весь финсчет ерзал на своих табуретах и подушечках , томимый любопытством. |
She sat upon cushions and leaned against an empty box and barrel, robe bespread, which formed a defence from the invading draughts. | Она восседала на принесенных из кареты подушках , прислонившись к пустому ящику и бочонку, устланным пледами и защищавшими ее от порывов сквозного ветра. |
It was only then that she turned her head and saw the face of the woman who had sagged back against the cushions . | Лишь после этого она повернула голову и увидела лицо женщины, откинувшейся на диванные подушки . |
Scarlett would have suffered agonies of humiliation at having friends find her without draperies and carpets and cushions and the proper number of chairs and teacups and spoons. | Скарлетт умерла бы от унижения, доведись ей принимать друзей в доме, где нет ни штор, ни ковров, ни диванных подушек , ни нужного количества стульев или чашек и ложек. |
She has thrown herself upon the floor and lies with her hair all wildly scattered and her face buried in the cushions of a couch. | Она бросилась на пол и лежит, зарывшись лицом в диванные подушки , а волосы ее разметались в беспорядке. |
Don’t you dare use my best cushions from the front room! | Только попробуй тронуть мои лучшие диванные подушки из гостиной! |
There aren’t enough cushions . | Здесь недостаточно диванных подушек … Поэтому я возьму это. |
Yeah, I found it between the sofa cushions . | Я нашла его в диване. |
With my brother we used to play that we had to step on cushions , pillows, clothes, ’cause touching the floor meant the end of the world. | Мы с моим братом раньше играли в эту игру, мы ходили по дивану, подушкам , одежде, а касание пола означало конец всему земному существованию. |
He sat stiffly on the vast, gray cushions of his limousine, not looking at her. | Он сидел, выпрямившись на больших серых подушках сидений своего лимузина, и не смотрел в её сторону. |
Scarlett gasped, and, for a moment, shrank back against the cushions . | Скарлетт ахнула и инстинктивно отстранилась от него, глубже уйдя в подушки сиденья. |
With our travel budget of whatever’s in the couch cushions , the world is our oyster. | А наш бюджет на путешествие — все что завалялось в диване, ведь мир — наша раковина. |
An ugly pile of rubble was lying on the road: pistons, cushions , springs. | На дороге валялась безобразная груда обломков: поршни, подушки , рессоры. |
To Tom Canty, half buried in his silken cushions , these sounds and this spectacle were a wonder unspeakably sublime and astonishing. | Для Тома Кенти, утопавшего в шелковых подушках , эти звуки и это зрелище были чудом, несказанно великолепным, поразительным. |
Formica table tops, plastic cushions and cups and saucers were all canary colour. | Столы, пластиковые подставки, чашки и блюдца — все сияло канареечным цветом. |
It sat among cushions and was surrounded with rag dolls and other toys in modern excess. | Она сидела меж подушек , на полу валялись тряпичные куклы и, как теперь принято в семьях, множество игрушек. |
Okay, so, the cushions are gonna be, like, rocks or tiny islands or lily pads that can take your weight. | Подушки — это, предположим, камни, или островки, или кувшинки, которые выдержат твой вес. |
Do you know I couldn’t sleep for a month because I got a dot of ink on one of the sofa cushions ? | А ты знаешь, что я месяц не спала потому что поставила чернильное пятно на диванной подушке ? |
I took off the slipcovers, had them dry-cleaned and then fumigated the cushions . | Я сняла чехлы, отдала их в химчистку, а после продизенфицировала подушки . |
Used to cut foam for furniture cushions . | Использовались для разрезания мебельных подушек . |
I’m gonna check my office for whoopee cushions . | Пойду, поищу в своем кабинете пукательные подушки . |
Horsehair curling out of seat cushions , fenders battered and hammered back. | Конский волос клочьями торчит из подушек , помятые, облупившиеся крылья. |
loved to buy scatter cushions and throw them around the gaff. | нравилось покупать декоративные подушки и разбрасывать их по дому в выходной. |
The hot tears welled into his eyes; he tore his hand away, and, flinging himself on the divan, he buried his face in the cushions , as though he was praying. | Горячие слезы подступили к глазам Дориана, он вырвал свою руку из руки Холлуорда и, упав на диван, спрятал лицо в подушки . |
See how this bit of celadon in the border is picked up by these cushions ? | Посмотрите, как эта часть цвета морской волны возвышается благодаря подушкам . |
He’s been carving up cushions before he even committed the robbery. | Он разрезал подушки до того как совершил ограбление. |
They tenderly lifted Toad into the motor-car and propped him up with soft cushions , and proceeded on their way. | Они осторожно подняли мистера Тоуда, и отнесли в машину, и подложили мягкие подушки , а потом двинулись дальше. |
Zoe sank back on the cushions and Rolling looked gloomily at her. | Роллинг невесело посмотрел на нее. |
In the confusion of cushions and quilts and curtains, at first few of the attendants’ blows found me. | Среди подушек , шестов и занавесей первые удары телохранителей почти не чувствовались мной. |
I am here, on my knees holding up these couch cushions as a symbol of my sorrow and regret. | Вот он я, стою на коленях держу эти диванные подушки как символ своих скорби и раскаяния. |
She glanced at the cushions , squashed out of shape. | Она с отчаянием посмотрела на мятые подушки . |








 .. so I’ll be the one to use it.
.. so I’ll be the one to use it.




cushion — перевод на русский в контексте, транскрипция кушен с английского на русский примеры
Bring the cups.
Ismael’s doing his cushion thing.
Not so loud.
Принеси чашки.
Исмаель укачивает подушку.
Не так громко.
Скопировать
Are you crazy?
I loved that cushion!
— I’ll get it.
Ты с ума сошел?
Я так любила эту подушку!
Я ее принесу.
Скопировать
— Not now…
I’ll get the cushion.
Jealous…
— Не сейчас. ..
..
Меня это смущает, лучше схожу за подушкой.
О, какая ревность…
Скопировать
sit on me with your cushion-butt?
It won’t hurt, ’cause it’s a cushion-butt.
Okay. If you could be the princess in any country, what country would it be?
Сядешь на меня своей жоподушкой?
Мне не будет больно, это же жоподушка.
Если бы ты могла быть принцессой в любой стране, ты бы какую выбрала?
Скопировать
I’m an evil mastermind.
Kelso, is there a cushion glued to my butt?
No. Not glued, superglued.
Я злобный гений.
Келсо, ты приклеил подушку к моей заднице?
Нет, не просто приклеил.
Скопировать
Please do.
Offer him a cushion.
No need, thank you.
Да пожалуйста.
Дайте ему подушечку.
Не нужно, спасибо.
Скопировать
Hey, hey, Kenny.
Cushion, bank manager.
Bank manager, bank manager!
Эй, Эй, Кенни.
Подушки, менеджер банка.
Bank manager, bank manager!
Скопировать
«Gift Day.» You idiots.
A whoopee cushion?
What are you, two?
«День подарков». Во идиоты.
Подушка-перделка?
Тебе что, 2 года?
Скопировать
What are you gonna do…
sit on me with your cushion-butt?
It won’t hurt, ’cause it’s a cushion-butt.
И что ты сделаешь?
Сядешь на меня своей жоподушкой?
Мне не будет больно, это же жоподушка.
Скопировать
Thanks for the tip.
Maybe I can use your body to cushion the blow.
HIGH PRIEST: I doubt my form would provide you much protection.
Спасибо за совет.
Может, я смогу воспользоваться твоим телом, чтобы смягчить падение.
Я сомневаюсь, что моя форма даст тебе много смягчения.
Скопировать
-Too late.
Here’s a cushion with a Chinese pattern.
25 öre.
— Слишком поздно.
Вот подушка с китайским рисунком.
25 крон.
Скопировать
After all, they are emotionally inexperienced, with only a few years … in which to store up the experiences which you and I take for granted.
If we gift them with a past, we create a cushion or a pillow for their emotions … then consequently
Memories.
Ведь они эмоционально неопытны, у них всего несколько лет для хранения опыта, который мы с вами воспринимаем, как должное.
Если мы одариваем их прошлым, то создаем некую подушку, амортизатор для их эмоций и тогда, как следствие, мы можем лучше их контролировать.
Воспоминания.
Скопировать
That chair’s uncomfortable.
I’ll bring you a cushion.
No, I’m fine.
У тебя стул неудобный.
Я принесу подушку.
Нет, мне удобно.
Скопировать
Knock yourself out.
Why does she want your seat cushion?
I don’t know why she brought it, I don’t know why she’s taking it.
ЧАРЛИ Забирай.
АЛАН Зачем ей твоя подушка?
ЧАРЛИ Не знаю, зачем она её принесла, и зачем забирает. Ну, ладно.
Скопировать
It turns out we’re not gonna need a babysitter tonight. No problem.
I’m gonna take this seat cushion, okay?
Knock yourself out.
И оказывается нам сегодня не нужна няня.
РОУЗ Хорошо. Тогда я заберу подушку для кресла?
ЧАРЛИ Забирай.
Скопировать
Leave him.
I don’t want him bleeding over my cushion.
— Here you are, Harry.
Оставьте его.
Не хочу, чтобы он заляпал кровью подушки.
— Вот возьми, Гарри.
Скопировать
They’re having themselves a little dance tomorrow night.
Enough we could take there for a small cushion for the Zelton job?
Sunday night?
Они будут танцевать завтра ночью.
Мы могли немного отдохнуть перед зелтонской работой.
В воскресенье вечером?
Скопировать
Very well! Please go up.
Light like a down cushion!
It is true.
Полезайте на спину.
Лёгкая, словно подушка!
Это точно.
Скопировать
— What is it?
Lannington was smothered to death with a cushion.
Two of them.
— Что это такое?
Леннингтон был задушен подушкой.
Двое из них.
Скопировать
There my darling will pray.
You must have a cushion.
Is this armchair to your liking?
Здесь, моя дорогая, Вы будете молиться.
У Вас будет подушечка.
Вам нравится это кресло?
Скопировать
Look at that!
Last time, a dog came and ripped my cushion.
What should I throw at you?
вы гляньте.
В тот раз собака мне подушку изгрызла.
Чем бы тебя обезвредить?
Скопировать
Oh, it is you, Sir.
bring him a cushion.
Don’t worry about that.
Здравствуйте, офицер.
Принеси ему подушку.
Не беспокойтесь об этом.
Скопировать
We’re still having security problems.
Dad, there is nothing to sleep on except a cushion on the floor.
We can get you a real bunk off the Enterprise.
У нас всё ещё есть проблемы с безопасностью.
Пап, тут неначем спать, кроме подушки на полу!
Мы можем достать тебе настоящую койку с Энтерпрайза.
Скопировать
Dad’s birthday is tomorrow… I hope he’s happy with it.
He should be… with a cushion and a record… he’ll love Paganini!
I like the haircut… now we almost look like brothers!
Завтра папин день рождения — Я надеюсь он будет счастлив.
Ему должно понравиться—с подушкой и записью… он будет любить Паганини!
Мне нравится стрижка — сейчас мы как братья!
Скопировать
BUT IF YOU DIDN’T MEAN TO STAY WITH ME, YOU’D BETTER NEVER HAVE COME.
THOSE THAT NEVER HAD A CUSHION DON’T MISS IT. NAY, NAY, AUNT!
NAY, NAY, AUNT!
I SAID I’D STAY ONLY FOR AS LONG AS I’D BE OF ANY COMFORT TO YOU. AND WHO SAID YOU’VE LEFT OFF
Если ты не хотела остаться со мной, лучше б не приезжала.
Кто не знал подушек, и без них выспится.
Нет, я же обещала остаться, пока смогу утешать вас.
Скопировать
I had a bath, I did not delay any more than ten minutes.
While returning, it had that in the cushion.
— Only two words: » To I halve it. «
Я принимал ванну.
Отсутствовал не больше десяти минут, а когда вернулся, нашёл вот это.
— Всего три слова: в 12 часов.
Скопировать
— Second question…
Someone of the house put the message in the cushion of the Waverly.
— Yes, it does sense.
— Пункт второй.
Кто-то из своих должен был подкинуть записку в комнату мистера Вэйверли.
— Да, согласен.
Скопировать
She just wants to try it.
But we can still date — like this is supposed to cushion the blow.
I got married so I could stop dating, so still dating is not a big incentive since the last thing you wanna do is date your wife, who should love you.
Просто попробовать.
Но мы можем встречаться. — Как будто это может смягчить удар.
Я женился, чтобы перестать встречаться, поэтому свидания — не стимул. Кому захочется назначать свидания собственной жене, которая тебя любит.
Скопировать
And, clear, for sale of work.
She knows, wanted, of certainty that this cushion is wet.
Allow me.
И благотворительную распродажу.
По-моему, подушка свалилась, дорогая.
Позволь мне.
Скопировать
Have a safe trip and have fun.
I can’t find the inflatable cushion.
It has to be here.
Отдыхайте. — Спасибо.
— Спасибо.
— Никак не найду надувную подушку. — Должна быть там.
Ищи лучше.
Скопировать
Показать еще
100 самых популярные английских слов на тему Дом и быт с переводом и произношением
Здесь мы собрали для вас 100 самых популярные английских слов на тему Дом и быт. В этот список слов-существительных вошли: части дома, помещения и предметы домашнего обихода, а также другие часто употребляемые слова на эту тему.
Все слова помимо русского перевода и фонетической транскрипции и имеют правильное произношение этих слов и упражнения для лучшего запоминания под списком, вы также можете пройти тест на знания лексики на эту тему.
- Слова из темы
- Учить слова
| Слово | Перевод | |
|---|---|---|
| 6157 |  mp3″>aerial mp3″>aerial
[ˈeərɪəl] ˈэариэл |
антенна |
| 2382 | apartment
[əˈpɑːtmənt] эˈпа:тмэнт |
квартира; апартамент |
| 10144 | armchair
[ˈɑːmˌtʃeər] ˈа:мˌчэар |
кресло; кабинетный |
| 6885 | balcony
[ˈbælkənɪ] ˈбэлкэни |
балкон |
| 4691 |  mp3″>barn mp3″>barn
[bɑːn] ба:н |
сарай; амбар |
| 4697 | basement
[ˈbeɪsmənt] ˈбэйсмэнт |
подвал; цокольный этаж; фундамент; основание; подвальный этаж |
| 3089 | bath
[bɑːθ] ба:сэ |
ванна; баня; купание; купать |
| 3844 | bathroom
[ˈbɑːθruːm] ˈба:сэру:м |
ванная комната; ванная; туалет |
| 802 |  mp3″>bed mp3″>bed
[bed] бэд |
постель; кровать |
| 17095 | bedclothes
[ˈbedkləʊðz] ˈбэдклоузз |
постельное белье |
| 2839 | bedroom
[ˈbedruːm] ˈбэдру:м |
спальня |
| 7820 | bedside
[ˈbedsaɪd] ˈбэдсайд |
прикроватный |
| 17224 |  mp3″>bedspread mp3″>bedspread
[ˈbedspred] ˈбэдспрэд |
постельное покрывало |
| 4460 | blanket
[ˈblæŋkɪt] ˈблэнкит |
одеяло; покров; защитный слой |
| 14136 | bookcase
[ˈbʊkkeɪs] ˈбуккэйс |
книжный шкаф |
| 15239 | bookshelf
[ˈbʊkʃelf] ˈбукшэлф |
книжная полка |
| 5936 |  mp3″>bucket mp3″>bucket
[ˈbʌkɪt] ˈбакит |
ведро; бадья |
| 2678 | cabinet
[ˈkæbɪnət] ˈкэбинэт |
кабинет |
| 5033 | carpet
[ˈkɑːpɪt] ˈка:пит |
ковер; покрытие |
| 3298 | ceiling
[ˈsiːlɪŋ] ˈси:лин |
потолок; перекрытие |
| 1434 |  mp3″>chair mp3″>chair
[tʃeər] чэар |
стул; кресло; председатель; кафедра; председательствовать |
| 7599 | chimney
[ˈtʃɪmnɪ] ˈчимни |
дымоход; камин; дымовая труба |
| 5201 | closet
[ˈklɒzɪt] ˈклозит |
чулан; стенной шкаф; уборная |
| 14340 | cornice
[ˈkɔː. ˈко:нис |
карниз; свес |
| 9410 | cot
[kɒt] кот |
детская кроватка; раскладушка |
| 4384 | cottage
[ˈkɒtɪdʒ] ˈкотидж |
коттедж; изба |
| 4831 | couch
[kaʊtʃ] кауч |
диван; кушетка; тахта; ложе |
| 5335 |  mp3″>courtyard mp3″>courtyard
[ˈkɔːtjɑːd] ˈко:тйа:д |
двор; внутренний двор |
| 1766 | cup
[kʌp] кап |
чашка; кубок; чаша; чашечка |
| 9160 | cupboard
[ˈkʌbəd] ˈкабэд |
шкаф; буфет; чулан |
| 4959 | curtain
[ˈkɜːtən] ˈкётэн |
занавес; завеса; занавеска |
| 8341 |  mp3″>cushion mp3″>cushion
[ˈkʊʃən] ˈкушэн |
подушка; смягчать |
| 2180 | desk
[desk] дэск |
стол; письменный стол; рабочий стол; регистратура; настольный |
| 13927 | dishwasher
[ˈdɪʃˌwɒʃər] ˈдишˌвошэр |
посудомоечная машина; посудомойка |
| 486 | door
[dɔːr] до:р |
дверь; дверной; дверца |
| 7075 |  mp3″>driveway mp3″>driveway
[ˈdraɪvweɪ] ˈдрайввэй |
дорога; проезд |
| 10504 | dryer
[ˈdraɪər] ˈдрайэр |
сушилка; фен |
| 5413 | elevator
[ˈelɪveɪtər] ˈэливэйтэр |
лифт; элеватор; подъемник |
| 13816 | faucet
[ˈfɔːsɪt] ˈфо:сит |
кран; вентиль; водопроводный кран |
| 3936 |  mp3″>fence mp3″>fence
[fens] фэнс |
забор; ограда; ограждение; загородка; изгородь; фехтовать |
| 6389 | fireplace
[ˈfaɪəpleɪs] ˈфайэплэйс |
камин; очаг |
| 1661 | flat
[flæt] флэт |
квартира; плоский |
| 879 | floor
[flɔːr] фло:р |
этаж; пол; земля |
| 11792 |  mp3″>flooring mp3″>flooring
[ˈflɔːrɪŋ] ˈфло:рин |
пол; настил |
| 17549 | footstool
[ˈfʊt.stuːl] ˈфутсту:л |
скамеечка для ног |
| 5092 | fork
[fɔːk] фо:к |
вилка; вилы; развилка; ответвление; камертон |
| 9890 | freezer
[ˈfriːzər] ˈфри:зэр |
морозилка |
| 11247 |  mp3″>fridge mp3″>fridge
[frɪdʒ] фридж |
холодильник |
| 2844 | furniture
[ˈfɜːnɪtʃər] ˈфёничэр |
мебель |
| 5041 | garage
[ˈɡærɑːʒ] ˈгэра:ж |
гараж |
| 1332 | garden
[ˈɡɑːdən] ˈга:дэн |
сад; огород; парк; садовый |
| 2134 |  mp3″>gate mp3″>gate
[ɡeɪt] гэйт |
ворота; затвор; калитка; шлюз |
| 16798 | guesthouse
[ˈɡesthaʊs] ˈгэстхаус |
дом для гостей; гостиница; пансион |
| 961 | hall
[hɔːl] хо:л |
зала; зал; холл; коридор |
| 194 | home
[həʊm] хоум |
дома; домой; дом; домашний; родной |
| 201 |  mp3″>house mp3″>house
[haʊs] хаус |
дом; жилище |
| 13385 | kennel
[ˈkenəl] ˈкэнэл |
конура; псарня; собачья конура |
| 8663 | kettle
[ˈketl] ˈкэтл |
чайник; котел; котелок |
| 497 | key
[kiː] ки: |
ключ; ключевой; клавиша; кнопка; основной |
| 1695 |  mp3″>kitchen mp3″>kitchen
[ˈkɪtʃɪn] ˈкичин |
кухня; кухонный |
| 1695 | kitchen
[ˈkɪtʃɪn] ˈкичин |
кухня; кухонный |
| 2990 | knife
[naɪf] найф |
нож; ножевой |
| 3898 | lamp
[læmp] лэмп |
лампа; светильник; фонарь; ламповый |
| 4988 |  mp3″>lawn mp3″>lawn
[lɔːn] ло:н |
газон; лужайка |
| 5689 | linen
[ˈlɪnɪn] ˈлинин |
белье; льняной; полотно; полотняный |
| 9506 | loft
[lɒft] лофт |
чердак; лофт; верхний этаж |
| 8204 | mattress
[ˈmætrəs] ˈмэтрэс |
матрац; тюфяк |
| 2179 |  mp3″>mirror mp3″>mirror
[ˈmɪrər] ˈмирэр |
зеркало; зеркальце; отражать |
| 10023 | napkin
[ˈnæpkɪn] ˈнэпкин |
салфетка; подгузник |
| 5603 | nursery
[ˈnɜːsərɪ] ˈнёсэри |
детская; ясли; детская комната; рассадник |
| 4436 | oven
[ˈʌvən] ˈавэн |
духовка; печь; духовой шкаф |
| 1843 |  mp3″>painting mp3″>painting
[ˈpeɪntɪŋ] ˈпэйнтин |
живопись; картина; покраска; роспись; окраска |
| 5581 | pillow
[ˈpɪləʊ] ˈпилоу |
подушка |
| 1517 | plate
[pleɪt] плэйт |
тарелка; плита; пластинка; лист |
| 7385 | quilt
[kwɪlt] квилт |
стеганое одеяло; стегать; подбивать ватой; простегивать |
| 2408 |  mp3″>roof mp3″>roof
[ruːf] ру:ф |
крыша; кровля |
| 309 | room
[ruːm] ру:м |
комната; номер; помещение; место |
| 7658 | scissors
[ˈsɪzəz] ˈсизэз |
ножницы |
| 12304 | shampoo
[ʃæmˈpuː] шэмˈпу: |
шампунь |
| 4033 |  mp3″>shelf mp3″>shelf
[ʃelf] шэлф |
полка; шельф |
| 4323 | shower
[ʃaʊər] шауэр |
душ; ливень; осыпать; принимать душ |
| 3820 | sink
[sɪŋk] синк |
раковина; потопить; слив; сток; тонуть; погружаться; топить |
| 5660 | sofa
[ˈsəʊfə] ˈсоуфэ |
диван; софа |
| 6065 |  mp3″>spoon mp3″>spoon
[spuːn] спу:н |
ложка |
| 6528 | staircase
[ˈsteəkeɪs] ˈстэакэйс |
лестница; лестничная клетка |
| 5992 | stool
[stuːl] сту:л |
стул; табурет; табуретка |
| 5470 | stove
[stəʊv] стоув |
плита; печь; печка |
| 206 |  mp3″>table mp3″>table
[ˈteɪbl] ˈтэйбл |
стол; таблица |
| 13665 | tablecloth
[ˈteɪblklɒθ] ˈтэйблклосэ |
скатерть |
| 4570 | tap
[tæp] тэп |
кран; отвод; ответвление; метчик |
| 5038 | teaspoon
[ˈtiːspuːn] ˈти:спу:н |
чайная ложка |
| 5193 |  mp3″>toilet mp3″>toilet
[ˈtɔɪlɪt] ˈтойлит |
туалет; ванная; туалетный |
| 12957 | toothpaste
[ˈtuːθpeɪst] ˈту:сэпэйст |
зубная паста |
| 5955 | towel
[taʊəl] тауэл |
полотенце |
| 6005 | trash
[træʃ] трэш |
мусор; хлам; дрянь; халтура; отбросы |
| 7672 |  mp3″>vase mp3″>vase
[vɑːz] ва:з |
ваза |
| 10307 | wallpaper
[ˈwɔːlˌpeɪpər] ˈво:лˌпэйпэр |
обои |
| 8735 | wardrobe
[ˈwɔːdrəʊb] ˈво:дроуб |
гардероб; платяной шкаф; гардеробная |
| 804 | window
[ˈwɪndəʊ] ˈвиндоу |
окно; окошко; оконный |
| 2872 |  mp3″>yard mp3″>yard
[jɑːd] йа:д |
двор; ярд; сад; парк |
 nɪs]
nɪs]Помогите проекту, поделитесь в соц.сетях
Запоминайте слова «тема Дом»
Упражнение № 2Слушате и запоминайте слова из раздела 100 популярных английских слов по теме Дом (House), для лучшего запоминания повторяте их в слух за диктором
перевод
АнглийскийРусский
lawn
[lɔːn]газон, лужайка
kitchen
[ˈkɪtʃɪn]кухня, кухонный
window
[ˈwɪndəʊ]окно, окошко, оконный
fork
[fɔːk]вилка, вилы, развилка, ответвление, камертон
garage
[ˈɡærɑːʒ]гараж
closet
[ˈklɒzɪt]чулан, стенной шкаф, уборная
bathroom
[ˈbɑːθruːm]ванная комната, ванная, туалет
wardrobe
[ˈwɔːdrəʊb]гардероб, платяной шкаф, гардеробная
hall
[hɔːl]зала, зал, холл, коридор
trash
[træʃ]мусор, хлам, дрянь, халтура, отбросы
floor
[flɔːr]этаж, пол, земля
blanket
[ˈblæŋkɪt]одеяло, покров, защитный слой
tablecloth
[ˈteɪblklɒθ]скатерть
lamp
[læmp]лампа, светильник, фонарь, ламповый
plate
[pleɪt]тарелка, плита, пластинка, лист
kitchen
[ˈkɪtʃɪn]кухня, кухонный
shelf
[ʃelf]полка, шельф
carpet
[ˈkɑːpɪt]ковер, покрытие
scissors
[ˈsɪzəz]ножницы
door
[dɔːr]дверь, дверной, дверца
cup
[kʌp]чашка, кубок, чаша, чашечка
oven
[ˈʌvən]духовка, печь, духовой шкаф
towel
[taʊəl]полотенце
cabinet
[ˈkæbɪnət]кабинет
furniture
[ˈfɜːnɪtʃər]мебель
toilet
[ˈtɔɪlɪt]туалет, ванная, туалетный
garden
[ˈɡɑːdən]сад, огород, парк, садовый
cornice
[ˈkɔː. nɪs]карниз, свес
nɪs]карниз, свес
bookcase
[ˈbʊkkeɪs]книжный шкаф
stove
[stəʊv]плита, печь, печка
kennel
[ˈkenəl]конура, псарня, собачья конура
teaspoon
[ˈtiːspuːn]чайная ложка
bookshelf
[ˈbʊkʃelf]книжная полка
roof
[ruːf]крыша, кровля
wallpaper
[ˈwɔːlˌpeɪpər]обои
toothpaste
[ˈtuːθpeɪst]зубная паста
napkin
[ˈnæpkɪn]салфетка, подгузник
home
[həʊm]дома, домой, дом, домашний, родной
bedclothes
[ˈbedkləʊðz]постельное белье
dishwasher
[ˈdɪʃˌwɒʃər]посудомоечная машина, посудомойка
loft
[lɒft]чердак, лофт, верхний этаж
cot
[kɒt]детская кроватка, раскладушка
painting
[ˈpeɪntɪŋ]живопись, картина, покраска, роспись, окраска
chair
[tʃeər]стул, кресло, председатель, кафедра, председательствовать
gate
[ɡeɪt]ворота, затвор, калитка, шлюз
mirror
[ˈmɪrər]зеркало, зеркальце, отражать
yard
[jɑːd]двор, ярд, сад, парк
nursery
[ˈnɜːsərɪ]детская, ясли, детская комната, рассадник
flooring
[ˈflɔːrɪŋ]пол, настил
guesthouse
[ˈɡesthaʊs] дом для гостей, гостиница, пансион
couch
[kaʊtʃ]диван, кушетка, тахта, ложе
faucet
[ˈfɔːsɪt]кран, вентиль, водопроводный кран
cottage
[ˈkɒtɪdʒ]коттедж, изба
pillow
[ˈpɪləʊ]подушка
house
[haʊs]дом, жилище
apartment
[əˈpɑːtmənt]квартира, апартамент
sink
[sɪŋk]раковина, потопить, слив, сток, тонуть, погружаться, топить
staircase
[ˈsteəkeɪs]лестница, лестничная клетка
fireplace
[ˈfaɪəpleɪs]камин, очаг
tap
[tæp]кран, отвод, ответвление, метчик
linen
[ˈlɪnɪn]белье, льняной, полотно, полотняный
ceiling
[ˈsiːlɪŋ]потолок, перекрытие
balcony
[ˈbælkənɪ]балкон
shower
[ʃaʊər]душ, ливень, осыпать, принимать душ
fence
[fens]забор, ограда, ограждение, загородка, изгородь, фехтовать
bedroom
[ˈbedruːm]спальня
flat
[flæt]квартира, плоский
driveway
[ˈdraɪvweɪ]дорога, проезд
table
[ˈteɪbl]стол, таблица
mattress
[ˈmætrəs]матрац, тюфяк
cushion
[ˈkʊʃən]подушка, смягчать
bath
[bɑːθ]ванна, баня, купание, купать
bed
[bed]постель, кровать
key
[kiː]ключ, ключевой, клавиша, кнопка, основной
barn
[bɑːn]сарай, амбар
armchair
[ˈɑːmˌtʃeər]кресло, кабинетный
kettle
[ˈketl]чайник, котел, котелок
chimney
[ˈtʃɪmnɪ]дымоход, камин, дымовая труба
bedspread
[ˈbedspred]постельное покрывало
shampoo
[ʃæmˈpuː]шампунь
knife
[naɪf]нож, ножевой
bucket
[ˈbʌkɪt]ведро, бадья
fridge
[frɪdʒ]холодильник
stool
[stuːl]стул, табурет, табуретка
dryer
[ˈdraɪər]сушилка, фен
bedside
[ˈbedsaɪd]прикроватный
vase
[vɑːz]ваза
spoon
[spuːn]ложка
elevator
[ˈelɪveɪtər]лифт, элеватор, подъемник
aerial
[ˈeərɪəl]антенна
footstool
[ˈfʊt. stuːl]скамеечка для ног
stuːl]скамеечка для ног
basement
[ˈbeɪsmənt]подвал, цокольный этаж, фундамент, основание, подвальный этаж
room
[ruːm]комната, номер, помещение, место
cupboard
[ˈkʌbəd]шкаф, буфет, чулан
courtyard
[ˈkɔːtjɑːd]двор, внутренний двор
sofa
[ˈsəʊfə]диван, софа
freezer
[ˈfriːzər]морозилка
desk
[desk]стол, письменный стол, рабочий стол, регистратура, настольный
curtain
[ˈkɜːtən]занавес, завеса, занавеска
quilt
[kwɪlt]стеганое одеяло, стегать, подбивать ватой, простегивать
Play Stop
Тест на знание слов из темы
Оцените слова из набора
4. 8
8
оценили: 27 чел.
Комментарии к набору слов
BB cream, CC cream, DD, Cushion… Фантастический мир тональных кремов
Telegram
8 (800) 707-66-08
+7 (499) 653-72-81
пн–пт 09:00 — 18:00
Скидки 10% по промокоду — blog
- ликбез
- тесты
- обзор
- подборки
- лайфхаки
- вопрос-ответ
- битва
- ингредиенты
- хочу/могу
21/03/2017
автор Администратор Главный
Привет! Сегодня у нас будет интересное и увлекательное путешествие в мир тональников Южной Кореи. «Тональники» — это условное кодовое слово, которое означает покрытие с маскирующими функциями, ведь если копнуть поглубже — то под ним скрывается как минимум пять видов таких покрытий.
Ваш опыт, как и мой, скорее всего, мог начаться с bb-крема. Ведь несколько лет назад это был единственный доступный и возможный вариант, и тогда он был революцией! Прелесть bb-крема я оценила тогда, когда он вкупе с другим корейским уходом практически привел мою кожу в состояние идеальной. Сначала он снизил жирность, а затем начал потихоньку снимать воспаления и стирать несовершенства — за год кожа стала совершенно другой. Я бы ни за что не поверила, что это возможно, если бы это не случилось со мной.
Что есть BB крем? Это аббревиатура «Blemish Balm» — бальзам против несовершенств.
Текстура у него может слегка различаться, но в целом для бибишек характерна густая консистенция и высокая кроющая способность — бб-крем может основательно убрать красноту, скрыть прыщи и даже синяки. Что в нем необычного? В том, что помимо макияжа, он выполняет мощную защитную и лечебную функцию — увлажняет, питает, витаминизирует — в зависимости от ингредиентов, которые туда добавили.
BB-кремы в большинстве своем имеют серый подтон и всего два оттенка — 21 и 23. Это значит, что он не будет придавать ни красноту, ни желтить вашу кожу, а даст бежевый здоровый цвет лица.
21й — для светлокожих девушек, и 23 — для кожи с легким загаром или оливковым подтоном. Но что точно стоит знать — bb-крем в течение пяти минут адаптируется к цвету вашей кожи, поэтому промахнуться с цветом очень сложно.
Совет: Если вам хочется сделать из своего бб-крема полупрозрачный флюид, или вы посчитали, что промахнулись с оттенком, просто смешайте порцию с 1-2 каплями косметического масла для лица — или специального, или кокосового — и наносите. А если делать это влажным бьютиблендером, вы еще больше снизите плотность
CC-крем был вторым шагом корейских производителей после бб-крема, однако все равно не обрел должной популярности. Что он представляет из себя? Это аббревиатура, которую сами корейцы расшифровывают двумя способами: Color Correcting (коррекция цвета) и Color & Care (цвет и забота). И то, и то фактически является правильным. Ведь сс-крем — это гораздо больше уходовых ингредиентов, и очень интересная консистенция — в белом креме заключены «капсулы», которые «раскрываются» при нанесении и превращают его в светло-бежевое покрытие. Однако, сколько бы я не использовала такого типа кремов, это история для светлокожих девушек.
И то, и то фактически является правильным. Ведь сс-крем — это гораздо больше уходовых ингредиентов, и очень интересная консистенция — в белом креме заключены «капсулы», которые «раскрываются» при нанесении и превращают его в светло-бежевое покрытие. Однако, сколько бы я не использовала такого типа кремов, это история для светлокожих девушек.
Любой легкий загар — и вы невеста Эдварда Каллена. Да и светлокожих некоторые cc-кремы выбеливают, как гейш. Тут необходимо изучать отзывы и смотреть свотчи. На что способен такой вид покрытия? На то, чтобы осветлить кожу. Дать ей гораздо больше увлажнения и ухода, чем бб-крем.
Совет: Как избежать отбеливания и обеспечить продвинутый уход? Просто смешать бб и сс кремы в пропорции 1:1 или 1:2 в зависимости от желаемого оттенка
DD-крем. Самый непризнанный, самый редкий и самый спорный крем, который уж точно не играет первую скрипку в оркестре корейских «тональников». Как только его не называют…И «dynamic do-all», и «daily defense». По сути производители подбираются под буквы свои определения. Интереснее всего получилось у марки APIEU, которая предложила концепцию «Dark Diminish». Это покрытие, которое под действием кислорода окисляется гораздо меньше, чем обычные тональники. Окисление — причина потемнения и тусклости любой поверхности, а тут получается — что до самого вечера свежий и светящийся вид. В целом, dd-кремы можно пересчитать по пальцам одной руки, и они ничем не отличаются по своему внешнему виду, консистенции, кроющим способностям от прародителя.
Как только его не называют…И «dynamic do-all», и «daily defense». По сути производители подбираются под буквы свои определения. Интереснее всего получилось у марки APIEU, которая предложила концепцию «Dark Diminish». Это покрытие, которое под действием кислорода окисляется гораздо меньше, чем обычные тональники. Окисление — причина потемнения и тусклости любой поверхности, а тут получается — что до самого вечера свежий и светящийся вид. В целом, dd-кремы можно пересчитать по пальцам одной руки, и они ничем не отличаются по своему внешнему виду, консистенции, кроющим способностям от прародителя.
Cushion — взрывная новинка последних лет, которая на крутом повороте обошла всех. Она просочилась на мировую арену, когда монстры Lancome выпустили такой же. По поводу названия до сих пор идут споры. Как правильно: кушн или кушон? На самом деле верного ответа нет, ведь произносится и пишется так, как слышится. А транскрипция — это где-то посередине между вариантами выше. Кушон — это средство, заключенное в пористую губку внутри кейса, и наносится он специальным спонжем, поверхность которого напоминает смесь шелка и резины. Невероятно нежная штучка, которая позволяет моментально нанести флюид похлопывающими движениями. И именно два слова тут принципиальны — ФЛЮИД и ПОХЛОПЫВАЮЩИМИ. Ведь средство гораздо более жидкое, чем прыдыдущие образцы. А при нанесении ни в коем случае не надо его размазывать по коже — его надо вдавливать.
Кушон — это средство, заключенное в пористую губку внутри кейса, и наносится он специальным спонжем, поверхность которого напоминает смесь шелка и резины. Невероятно нежная штучка, которая позволяет моментально нанести флюид похлопывающими движениями. И именно два слова тут принципиальны — ФЛЮИД и ПОХЛОПЫВАЮЩИМИ. Ведь средство гораздо более жидкое, чем прыдыдущие образцы. А при нанесении ни в коем случае не надо его размазывать по коже — его надо вдавливать.
К недостаткам кушонов можно отнести их бодрый расход (3 месяца — это предел) и то, что он может не менее бодро съедаться кожей, теряя кроющие способности. Но не у всех и не всегда. К достоинствам — все остальное! Нежное шелковое покрытие, возможность наслаивать, феноменальную маскировку кожи при легкой текстуре, наличие рефиллов, эффектный дизайн упаковки… нужно ли продолжать? Кушон по праву — король нынешнего времени. Он объединил в себе все лучшее, что было в bb, cc, dd кремах и вывел это на новый уровень.
Есть у него и «клоны», которые, строго говоря, кушонами не являются. Например, так называемые BB cushions, которые представляют собой знакомый нам густой бб крем в губке. Или просто BB pact / tension pact — фактически крем-пудра, упакованные в кейс. Поэтому обращайте внимание на надписи перед тем, как покупать.
Например, так называемые BB cushions, которые представляют собой знакомый нам густой бб крем в губке. Или просто BB pact / tension pact — фактически крем-пудра, упакованные в кейс. Поэтому обращайте внимание на надписи перед тем, как покупать.
Совет: Не реже, чем раз в три дня необходимо стирать спонж. Иначе рискуете потерять инструмент, ведь он падёт смертью храбрых, не отмывшись и поселив в себе колонию бактерий. Как эффективно отмыть спонж? Кладете в пластиковый пакет с застежкой zip-lock, наливаете внутрь немного воды, гидрофильного масла или мыльной пенки, и начинаете прокатывать поверхность бутылкой или скалкой. Вы как будто выдавливаете из спонжа все скопившееся в нем покрытие. После того, как проделаете это пару-тройку раз, можете прополоскать — как новый он не будет, но чистота гарантирована. Ни в коем случае не пытайтесь теребить его руками — разорвется
Foundation — он же классический тональный крем. Да, такие в Корее тоже бывают! И как раз в этом случае они ничем не отличаются от своих европейских собратьев, разве что объемом. На объем корейцы не скупятся: упаковки по 40-50 мл вместо стандартных 30 мл — это нормально и правильно.
На объем корейцы не скупятся: упаковки по 40-50 мл вместо стандартных 30 мл — это нормально и правильно.
Стик (он же bb-стик, bc-стик, cc-стик, или просто — Stick) — это тон-карандаш в твердой форме на жировой основе. Обычно он имеет плотное покрытие, много смягчающих масел, и оснащен спецэффектами. Например, сияющим хайлайтером в центре стика. При смешении получается интересный вариант влажной кожи. И хотя многие стики маркируются, как подходящие для жирной кожи, их реальная участь — уход и питание нормальной и сухой кожи. Если ваше лицо блестит, как в Масленицу, то стиками вам лучше не баловаться.
Так какой же «тональник» вам нужен?
Если вам нужна плотная текстура и хорошая маскировка — для вас bb-крем
Если вы хотите легких текстур и всех плюсов покрытия, и готовы мириться с расходом — кушон будет вашим лучшим другом
CC-крем — для тех, кто хочет осветлить и выровнять кожу, и не чувствовать дискомфорта в течение дня.
Стик подойдет вам, если кожа сухая или требует питания
Ну а foundation — для тех, кто привык к европейским тональником, но хочет начать переход к корейским покрытиям.
Лучшие в своем роде:
BB крем A’PIEU Natural Flawless Cover BB Cream SPF30/PA+++
ВВ крем Holika Holika Petit BB Cream
BB крем Missha M Signature Real Complete BB Cream
СС крем Etude House CC Cream #1 Silky
СС крем SKIN79 Complete CC Cream Control
СС крем Secret Key Let Me Know CC Cream
Кушон Missha M Magic Cushion Moisture
Кушон Holika Holika Gudetama Lazy & Easy Face 2 Change Photo Ready Cushion BB SPF50/PA+++
Крем-пудра Enprani Dear by Malang Pact-dation
Тональный крем Enprani Delicate Radiance Foundation
cushion сленг | cushion перевод, cushion значение
cushion
- En [ ˈkʊʃən]
- Us [ ]
Значение cushion в русском языке
-
0 a cloth bag filled with something soft that you sit on or lean against to make you comfortable — диванная подушка
-
1 something that protects you from possible problems — защита
-
Overseas savings provide a cushion against tax rises at home.

-
-
2 to reduce the bad effects of something — смягчать
-
Various attempts were made to cushion the impact of unemployment.
-
-
3 to protect something, especially part of the body, with something soft — смягчать
-
Soft grass cushioned his fall.
-

Дополнительные определения cushion
Примеры cushion
-
They have cushions that vary from short and squat, to more elongate and obovate.
-
In squatter cushions, the steep and narrow area surrounding the scar shows no ornamentation.
-
At some clubs, indeed, the dancers brought tapestry cushions embroidered with their names.
-
There are cushions made out of materials appropriate to the nature and technology theme and you can almost shut yourself in.
-
No specimen showed intimal cushions but little diffuse intimal thickening was present in 10 of 14 cases.
-
By adding to its rigidity it supports the closing mechanism by the endocardial cushions as long as valves have not yet developed.
-
Older staff facing structural and technological change in the workplace could bow out ‘ gracefully ‘, cushioned by lump sum payments and secure pensions.
-
However, these studies have been conducted on a variety of animal models, and without comparison between the two types of cushions.


Дополнительные примеры cushion
Тенденции cushion
Переводы cushion на другие языки
-
中文繁体
墊子, 座墊, 靠墊…
More -
中文简体
垫子, 座垫, 靠垫…
More -
Español
cojín, amortiguar, suavizar…
More -
Português
almofada…
More -
日本語
クッション…
More -
Türk dili
yastık, minder, muhtemel sorunlardan koruyan…
More -
Français
amortir, protéger contre, coussin [masculine]…
More -
Catalan
coixí…
More
Другие переводы cushion
Просматривать
- curtail
- curtain
- curtsey
- curve
- cushy
- custard
- custodial
- custodian
НОВЫЕ СЛОВА
European
May 10, 2021
Подробнее
СЛОВО ДНЯ
Shimmer
May 10, 2021
Подробнее
Навигация по сайту
- Дом
- Перевести
- Фонетический символ
- Словарь
- Дом
- Слова на букву K
- Слова на букву A
- Слова на букву G
- Перевести
- Фонетический символ
Отказ от ответственности: если ресурсы, включенные в этот сайт, нарушают ваши права, отправьте нам электронное письмо, мы своевременно удалим контент, нарушающий авторские права, спасибо за сотрудничество!
Copyright © 2011-2020 All Rights Reserved.
Series 02 Episode 16 – The Cushion Saturation
Сцена: Пейнтбольный полигон.
Говард: Это было близко.
Радж: Боже, как я люблю запах шаров с краской по утрам.
Говард: Да, все равно смешно, Радж.
Леонард: Нам никак не добраться до хребта. Химический факультет нас полностью отрезал.
Говард: А как же русло ручья?
Шелдон: Отдел фармакологии контролирует это, и все они накачаны экспериментальными стероидами.
Радж: Что ж, все, мы обречены.
Шелдон: Я думаю, пришло время признать, что мы расплачиваемся за то, что некоторые из нас не смогли посетить мое совещание по стратегии пейнтбола на физическом факультете.
Говард: Я же говорил, что у моей мамы сосудистые звездочки, мне пришлось отвезти ее в лазерную клинику.
Шелдон: А я говорил тебе, что хочу увидеть справку от врача.
Радж: Нам нужен план. Как насчет операции «Молот богов»?
Леонард: Я забыл. Какой из них Молот богов?
Радж: Мы прячемся за мусорными баками на стоянке и устраиваем засады на людей, когда они приходят пописать.
Говард: Не ходи. Мусорные контейнеры находятся глубоко на территории Департамента астрономии.
Леонард: Нет, это не проблема, Венера днем не спит. Они, наверное, просто смотрят в небо.
Шелдон: Хорошо, сейчас нам нужно тактическое отступление. Вы видели эпизод «Звездных врат», где они оказались на планете с культурой, основанной на земных Афинах и Спарте? Не важно. Леонард, Радж и я собираемся вырваться за дверь и убежать, Ховард нас прикроет.
Говард: Почему бы мне не убежать, а ты меня прикроешь?
Шелдон: Потому что ты предпочел вены своей матери победе. На три. Раз, два, три, вперед!
Говард: Я должен был взять ее! Погода почти в купальниках! (Другой игрок врывается в дверь)
Я сдаюсь! Не стреляйте! Они пошли туда!
Лесли: Говард, я в твоей команде.
Говард: О, Лесли, слава богу.
Лесли: Где остальная часть твоего отряда?
Говард: Ах, они бросили меня здесь умирать. Что насчет твоего?
Лесли: Все мертвы.
Говард: Извини.
Лесли: Не надо. Это был дружественный огонь. Они просто не слушали.
Говард: Что ж, мы окружены, так что, думаю, нам нечего делать, кроме как ждать, пока нас схватят или убьют.
Лесли: Хм, это самое худшее — ожидание.
Говард: Все время зная, что там есть шарик с краской с вашим именем.
Лесли: Да, большой мокрый шар смерти. Что-то заставляет вас чувствовать себя более живым, не так ли?
Говард: Вроде как.
Лесли: Я говорю, что мы ценим каждое мгновение.
Говард: Согласен. Как именно мы это делаем? (Лесли целует его)
Леонард (голос) : Говард, почему ты нас не прикрываешь? Нас тут зарежут!
Говард: Война — это ад.
Последовательность кредитов.
Сцена: Университетская столовая.
Радж: Ребята, вы видели новую бюджетную записку, которая вышла сегодня утром?
Леонард: Да, еще сокращения.
Шелдон: Недопустимо. Меня сбивает с толку, почему они просто не отпустят некоторых из вас, чтобы были деньги на мои исследования.
Меня сбивает с толку, почему они просто не отпустят некоторых из вас, чтобы были деньги на мои исследования.
Леонард: Знаешь, что меня сбивает с толку, Шелдон?
Шелдон: Основываясь на вашей успеваемости, могу предположить, что угодно.
Лесли: Привет, ребята.
Леонард: Привет, Лесли.
Говард: Привет.
Лесли: Говард, я получил разрешение на быстрый прототип, который ты хотел.
Говард: Отлично, Лесли. Спасибо.
Лесли: Ты почеши мне спину, я почешу тебе. Rawr.
Радж: Что это было?
Говард: Ну, ничего страшного. Они дали Лесли контроль над неограниченным грантом.
Леонард: Да, ладно, но что за почесывание спины и мяуканье!
Шелдон: Я полагаю, что метафора «чесание спины» обычно описывает ситуацию «услуга за услугу», когда одна сторона предоставляет товары или услуги другой стороне в качестве компенсации за аналогичное действие.
Леонард: Спасибо.
Шелдон: Мроу, мне показалось, что это африканская циветта.
Леонард: Ты закончил?
Шелдон: Нет. Несмотря на название, циветта не настоящая кошка. Теперь я закончил.
Радж: Знаешь, о чем я думаю? Я думаю, Ховард не придумывал метафору, чесающую спину. Я думаю, что были какие-то настоящие царапины.
Леонард: Как насчет этого, Говард?
Говард: Хорошо, я не хотел ничего говорить, потому что я знаю, что у вас с Лесли есть небольшая история.
Леонард: Меня это не волнует.
Говард: Отлично, потому что мне очень хотелось кое-что сказать.
Леонард: Ты и Лесли?
Говард: В пейнтбольном сарае! Дважды!
Шелдон: Поэтому вы не прикрыли наш побег и не позволили нам перерезаться, как животным?
Говард: О, да, прости.
Шелдон: Дорогой мой, неисполнение служебных обязанностей перед лицом врага карается военным трибуналом.
Говард: Военный трибунал, военный трибунал, Лесли Уинкль — пятая девушка, с которой я когда-либо занимался сексом. В смысле бесплатно.
Радж: К тому же у тебя есть быстрый прототип. Это дорогое оборудование, чувак.
Это дорогое оборудование, чувак.
Леонард: А у остальных урезан бюджет до минимума.
Говард: Хорошо, один из способов взглянуть на это так: я получаю новое оборудование, а ты нет, и это несправедливо. Но лучше посмотреть на это так: я занимаюсь сексом, а ты нет, и это восхитительно.
Сцена: Квартира. Леонард смотрит на ноутбук Пенни.
Пенни: Все замерло. Я не знаю, что случилось.
Леонард: Успокойся. Мы с этим разберемся.
Пенни: Как мне успокоиться? Я потеряю всю свою корзину. Три часа выбирания обуви просто к черту.
Шелдон: Привет, Пенни.
Пенни: Привет, Шелдон.
Шелдон: Ты на моем месте.
Пенни: Ты собираешься сидеть здесь?
Шелдон: Нет, я иду в магазин комиксов.
Пенни: Тогда какая разница?
Шелдон: Какая разница?
Леонард: Поехали.
Шелдон: Это мое место. В постоянно меняющемся мире это единственная точка непротиворечивости. Если бы моя жизнь была выражена как функция в четырехмерной декартовой системе координат, это место в тот момент, когда я впервые сел на него, было бы ноль-ноль-ноль-ноль.
Пенни: Что?
Леонард: Не садись на его место.
Пенни: Хорошо. (Ходит) Счастлив?
Шелдон: Я не несчастен.
Пенни: Боже, я люблю его, но он настоящий чудак.
Леонард: Хорошо, должно сработать.
Пенни: О, спасибо. Вы спасатель.
Леонард: Ты заказал много обуви.
Пенни: Да, знаешь, самое печальное, что на самом деле это не так. О, это одно из тех пейнтбольных ружей?
Леонард: Ага. Вы должны выйти с нами как-нибудь.
Пенни: О, нет, спасибо. Я из Небраски. Когда мы стреляем в вещи, это потому, что мы хотим их съесть или заставить их оставить наших парней в покое. (Пистолет стреляет и стреляет тремя шариками краски прямо в точку Шелдона.)
Сцена: спальня Говарда.
Лесли: Боже, твое сердце колотится. Должно быть, я действительно завел тебя.
Говард: Ну, отчасти это ты, отчасти моя преходящая идиопатическая аритмия.
Лесли: Секси.
Говард: Могу ли я предположить, что вы тоже нашли опыт…
Лесли: Удовлетворительно?
Говард: Это было не совсем то слово, которое я искал, но, конечно, я сделаю это.
Лесли: Эй, тебе нравится мой прототип?
Говард: О, это здорово. Все в инженерном отделе рвут душу.
Лесли: Разве это не мило, когда твоя удача делает других несчастными?
Говард: Знаете, большинство людей этого не понимают.
Мать Говарда (голос) : Говард, я дома!
Говард: О, отлично.
Мать Говарда (голос) : Книжный клуб отменили! Эта штука на шее Филлис снова открылась!
Говард: Я занят, ма!
Мать Говарда (голос) : Слишком занята, чтобы помочь маме с молнией?
Говард: Не входи, мам!
Мать Говарда (голос) : Почему бы и нет?
Лесли: У него компания!
Говард: А, это аритмия.
Мать Говарда (голос) : Она еврейка?
Говард: Ты еврей?
Лесли: Нет.
Говард: Да!
Мать Говарда (голос) : Хорошо, дети, веселитесь! Используйте защиту!
Сцена: Квартира. Леонард пытается смыть краску с сиденья Шелдона.
Пенни: Думаешь, он заметит?
Леонард: Шанс есть.
Пенни: Что будем делать?
Леонард: Мы? Нет, нет, нет, у тебя был шанс быть нами уже полтора года. Прямо сейчас ты это ты, и ты облажался.
Пенни: Почему мы должны говорить ему, что это сделала я?
Леонард: Ну, мы не собираемся говорить ему, что это сделал я.
Пенни: Ладно, ладно, как насчет этого? Мы говорим ему, что кто-то вломился.
Леонард: Просто чтобы подстрелить диван из пейнтбольного ружья?
Пенни: Прости, я куплюсь. Все эти люди на наркотиках.
Леонард: Мы могли бы сказать ему, что они хотели, чтобы диван держался подальше от их бойфренда.
Пенни: Хорошо, хорошо. Ну, а если просто перевернуть? Там. Выглядит хорошо, верно?
Леонард: Мм, отпечаток задницы. Нет заметного отпечатка приклада.
Пенни: Да ладно. (Сидит и ерзает) Вот, отпечаток приклада.
Леонард: Он слишком маленький и слишком совершенный.
Пенни: Спасибо.
Леонард: Пожалуйста.
Пенни (входит Шелдон) : Тсс! Веди себя нормально.
Леонард: Шелдон! Как магазин комиксов?
Шелдон: Хорошо. Вышел новый номер Flash.
Леонард: Отлично, отлично. Вы прошли весь путь? Немного прохладно.
Шелдон: Меня подобрал Кутраппали.
Леонард: Разве это не потрясающе? Он такой хороший друг. Знаете, что самое лучшее в друзьях?
Шелдон: Они не болтают без умолку без особой причины.
Леонард: Нет, нет, друзья прощают мелочи.
Пенни: Знаешь, мне нужно пойти домой и вымыть голову.
Леонард: Не смей, мисси.
Шелдон: Привет, самый быстрый человек. Хочешь увидеть, как я прочитаю весь твой комикс? Хотите увидеть это снова? Что-то не так.
Леонард: Что ты имеешь в виду?
Шелдон: Не уверен. Это неправильно.
Леонард: Я не понимаю, о чем ты говоришь. Ах это. Пенни так и сделала.
Сцена: То же самое. Все едят. Шелдон сидит на спинке стула Леонарда. Он продолжает издавать стонущие звуки. Сказав «Извините», он пытается вернуться на свое место, но подушки там нет. Он пытается взгромоздиться на руку в разных положениях.
Он продолжает издавать стонущие звуки. Сказав «Извините», он пытается вернуться на свое место, но подушки там нет. Он пытается взгромоздиться на руку в разных положениях.
Леонард: Почему бы тебе просто не поесть в кресле?
Шелдон: Почему бы мне просто не поесть в кресле?
Пенни: Поехали.
Шелдон: Это мое рабочее кресло, там я работаю. Я не ем в кресле за столом и не работаю на своем месте. Я работаю в своем рабочем кресле и ем на своем месте.
Леонард: Чудак.
Говард: Знаешь, здесь есть очевидное решение. (Раджу) Вставай. (Радж делает это. Ховард передвигает свою подушку на место Шелдона) Вот. Задача решена. (Радж шепчет ему) Никого не волнует, где ты будешь сидеть. Вы не сумасшедший.
Шелдон: Извините, проблема не решена. Если бы вам случайно ампутировали голову, а вместо нее мы пересадили бы голову собаки, это было бы «решением проблемы»?
Леонард: Если бы это была твоя голова, так бы и было.
Пенни: Шелдон, мне очень, очень жаль, но это только на неделю. Ты не можешь быть немного гибким? Да извини. Я действительно не думал об этом.
Шелдон: Вы утверждаете, что это будет через неделю, но я не верю в вашу химчистку.
Пенни: Почему бы и нет?
Шелдон: Ты заметил табличку на его прилавке? Он не работает в химчистке на полную ставку. Он также делает ключи.
Леонард: О, ради всего святого, Шелдон.
Шелдон: Фокусировка важна. Был ли Майкл Дебейки организатором свадеб между пересадками сердца? Подрабатывал ли Александр Флеминг парикмахером? «Спасибо за открытие пенициллина, а теперь как насчет того, чтобы попробовать начес?»
Говард (после того, как звонит его телефон) : О, похоже, сегодня вечером у меня будет секс. Эй, детка…
Пенни: Его правая рука зовет его?
Леонард: Нет, это Лесли Винкль. Это долгая история.
Говард: Я заеду за тобой через десять минут. Господа, прощайте.
Леонард: Я думал, сегодня вечером мы будем играть в Halo.
Говард: Что мне делать, Леонард? Там есть женщина, которая хочет заняться со мной сексом. Вы понимаете, верно?
Пенни: Нет. Вовсе нет.
Говард: Тем не менее, я должен уйти. Кстати, я тебе говорил? Лесли потянула за кое-какие ниточки и отправила меня в исследовательскую поездку в Женеву, чтобы проверить суперколлайдер CERN.
Леонард: Это несправедливо. Вы даже не физик.
Говард: Хорошо, есть два взгляда на это…
Леонард: Уходи.
Говард: Пока.
Сцена: спальня Говарда.
Лесли: Тебе становится лучше.
Говард: Спасибо. Мне помогает, когда я тренируюсь с настоящей женщиной.
Лесли: Эй, послушай. В субботу моя сестра выходит замуж. Я… я хочу, чтобы ты пошел со мной. Это черный галстук.
Говард: Да уж, я бы не хотел.
Лесли: Почему бы и нет?
Говард: Когда я хожу на свадьбы, я обманываю коренастых подружек невесты. Я не знаю, что я буду делать со свиданием.
Лесли: О, хорошо. Я понимаю.
Я понимаю.
Говард: Спасибо.
Лесли: Эй, я очень сожалею о поездке в Женеву.
Говард: Что насчет этого?
Лесли: О, разве ты не слышал? Мне пришлось сократить количество людей, и вы не попали в этот список.
Говард: Когда это случилось?
Лесли: Около 12 секунд назад.
Говард: Ну, подожди. Вы хотите сказать, что если я не пойду на свадьбу, я не смогу поехать в Женеву?
Лесли: Вообще-то я стараюсь этого не говорить.
Говард: Ладно, извини, но мне от этого немного не по себе.
Лесли: Как так?
Говард: Потому что ты как будто контролируешь меня с помощью нового оборудования и исследовательских поездок.
Лесли: Ну, если бы я не контролировала тебя с помощью нового оборудования и исследовательских поездок, мне было бы неудобно.
Говард: Как так?
Лесли: Потому что тогда у нас были бы настоящие отношения с чувствами и прочим дерьмом.
Говард: Итак, суть в том, что я просто купленная и оплаченная секс-игрушка.
Лесли: Нет. Нет, совсем нет. Ты тоже леденец. Так? Что вы думаете?
Говард: Эй, мам, ты должна взять мне напрокат смокинг!
Мать Говарда (голос) : Прямо сейчас? Какой у тебя там секс?
Сцена: Квартира
Леонард: Почему ты присел?
Шелдон: Это мое место. Где еще мне присесть?
Леонард: Не знаю. Техас?
Пенни (приходит с подушкой) : Вот, пожалуйста. Только что из химчистки, как новый.
Леонард: Правда? Большой. Шелдон, смотри. Как новый.
Шелдон: Я очень сомневаюсь, что от того производителя ключей.
Пенни: Ну же, Шелдон. Просто попробуйте.
Шелдон: Хорошо.
Пенни: Вот, красиво и удобно уютно. Ноль, ноль, ноль.
Шелдон: Еще один ноль. Вы забыли параметр времени.
Пенни: Садись на чертов диван.
Шелдон: Нет.
Пенни: Что ты имеешь в виду, нет? Что с этим не так?
Леонард: Ничего, с ним что-то не так.
Пенни: Точно такой же. ..
..
Леонард: Пенни, Пенни, кажется, я знаю, что делать. Шелдон, у меня плохие новости.
Шелдон: Еще?
Леонард: Боюсь, что да. Помнишь цыпленка с кешью, который я приношу тебе по понедельникам?
Шелдон: Да. Из Сычуаньского дворца.
Леонард: Сычуаньский дворец закрылся два года назад.
Шелдон: Что? Откуда взялся мой цыпленок с кешью?
Леонард: Золотой дракон.
Шелдон: Нет. Нет, это неправильно. Наша еда всегда поставляется в контейнерах Szechuan Palace.
Леонард: Да, но до того, как они закрылись, я купил 4000 контейнеров. Я храню их в багажнике своей машины.
Шелдон: Но. О, это все меняет.
Леонард: Я подумал, что это может отвлечь его от подушки.
Шелдон: Что реально? Что не так? Как я могу знать?
Пенни: Ты это выдумал, да?
Леонард: О, Боже, как бы я хотел.
Шелдон: Леонард?
Леонард: Да, приятель?
Шелдон: Мне все еще не нравится эта подушка.
Сцена: Пейнтбольный полигон.
Леонард: Ладно, во-первых, Пенни, спасибо, что пришла.
Пенни: Спасибо за туфли.
Леонард: Пенни отличный стрелок. Думаю, у нас есть реальный шанс победить на этой неделе.
Говард: Какой план?
Леонард: Хорошо. Теперь мы все выбегаем. Шелдон и я срежем налево за этими деревьями. Радж, Ховард и Лесли стоят справа за камнями. Тогда у всех нас будет прекрасный вид, как Пенни выбегает и убивает всех остальных в поле зрения.
Шелдон: Хорошо, только одно, прежде чем мы начнем.
Леонард: Что такое, Шелдон? (Шелдон стреляет в Пенни)
Пенни: Какого черта?
Шелдон: Это для моей подушки.
Леонард: Шелдон, Пенни была нашей единственной надеждой.
Шелдон: Прости, Леонард. Но месть — это блюдо, которое лучше всего подавать холодным.
Пенни: К черту. (Стреляет в Шелдона)
Шелдон: Она не может стрелять в меня. Она мертва.
Леонард: Он прав. Вы не можете. (Стреляет в Шелдона)
Шелдон: Ну, если мы собираемся погрузиться в анархию (Стреляет в Леонарда)
Говард: Хорошо, увидимся.
Леонард: Куда ты идешь?
Лесли: Сдаться, потом Денни.
Нравится:
Нравится Загрузка…
Транскрипция наушников ACC0233 | Philips
Наушники для транскрипции ACC0233 | ФилипсНаушники для транскрипции
ACC0233Найти похожие продукты
- Обзор
- Характеристики
- Поддерживать
Непревзойденное качество звука
с наушниками Philips для транскрипции
Легкие наушники Y-style обеспечивают превосходное качество звука, оснащены мягкими амбушюрами для удобства ношения и подвесным кронштейном, с помощью которого гарнитура крепится к монитору.
Купить
в вашем любимом магазине
Купить
в вашем любимом магазине
Найдите местного продавца
Найдите местного продавца
Особенности
Профессиональная гарнитура, обеспечивающая концентрированную транскрипцию в загруженном офисе
Профессиональная гарнитура, обеспечивающая концентрированную транскрипцию в оживленных офисных условиях.
Аксессуары Philips помогут вам максимально эффективно использовать устройства для диктовки Philips. Они гарантируют оптимальные результаты и гарантируют долгий срок службы вашей продукции Philips.
Неодимовый магнит повышает чувствительность и качество звука
Неодимовый магнит повышает чувствительность и качество звука
Неодим — лучший материал для создания сильного магнитного поля, повышающего чувствительность и общее качество звука.
Легкая конструкция повышает комфорт при длительном использовании
Легкая конструкция повышает комфорт при длительном использовании
Прочные и легкие качественные материалы повышают комфорт при длительном ношении.
Подбородочная подставка для всех ушей для большего комфорта
Подбородочная подкладка для всех ушей для большего комфорта
Подбородочная посадка для большего комфорта.
Кабель длиной 3 метра/10 футов для свободы передвижения по рабочему столу
Кабель длиной 3 метра/10 футов для свободы передвижения по рабочему столу
Кабель длиной 3 метра/10 футов дает вам свободу передвижения по рабочему столу, не снимая гарнитуру.
Сопутствующие товары и аксессуары
Введите серийный номер программного обеспечения
Программа распознавания речи VoiceTracer работает только с аудиорекордерами Philips VoiceTracer. Чтобы загрузить установочный файл, введите серийный номер, затем нажмите Скачать .
Чтобы загрузить установочный файл, введите серийный номер, затем нажмите Скачать .
Найдите свой серийный номер
Серийный номер можно найти в кратком руководстве на упаковке продукта.
Загрузка…
Если загрузка не начинается, нажмите здесь, чтобы повторить попытку.
Звук- Акустическая система:
- открытый
- Тип магнита:
- неодимовый
- Диафрагма:
- Звуковая катушка 7 PET 64 купольная0672
- copper
- Sound quality:
- optimized for voice reproduction
- Impedance:
- 32 Ohm
- Maximum power input:
- 10 mW
- Sensitivity:
- 103 dB
- Speaker diameter:
- 14 mm
- Тип:
- динамический
- Система:
- Одиночная звуковая система
- Кабельное соединение:
- one-sided
- Cable length:
- 3 m/10 ft
- Connector:
- 3. 5 mm stereo, right angled
- Type of cable:
- copper
 5 mm stereo, right angled
5 mm stereo, right angled
- Headphones
- Подвесной кронштейн
- Клипса для одного уха
- Информация WEEE
- Все настольные системы диктовки и транскрипции Philips
- Philips Pocket Memo Digital Voice Decorder
- Philips Pocket Memo Mecing Decorder
- СОБСТВЕННЫЙ КОМПЛЕКТ 2011/65/EU (ROHS)
- .
- Размеры изделия:
- 230 мм × 130 мм × 25 мм/9 дюймов × 5 дюймов × 1 дюйм
- USB-аудиоадаптер Philips LFH9034
Материалы по маркетингу и продажам
- Брошюра на английском языке (222 КБ)
Регистрация
У вас есть этот продукт?
Следите за гарантийным покрытием вашего продукта
Получите легкий доступ к гарантийному покрытию продукта
Получите право на возврат денег, подарки и специальные предложения
Зарегистрируйте свой продукт
Гарантия
Более подробную информацию о Гарантия Филипс.
Гарантийный документ
Где купить
Найти местного продавца
Рекомендуемые розничные продавцы
AAA Price.com, Inc.
Посетить сайт >
DictationOne (DBA YB Sales)
Посетить сайт >
Цифровые голосовые системы/DictationStore
Посетить сайт >
Исполнительные системы связи
Посетить сайт >
Dolbey Systems, Inc. / TranscriptionGear
Посетить сайт >
dictationproducts. com
com
Посетить сайт >
Контакт Поиск магазина
Новостная рассылка
Подпишитесь и будьте в курсе последних новостей от нас
О фонетических подушках
Подход фонетической подушки к речи отражает мою убежденность в том, что актеры должны быть чувствительными людьми, которые страстно, проницательно, интуитивно воплощают в жизнь слова драматурга, направляя действие пьесы и заставляя слушателя узнавать и сопереживать. Я нахожу письменные фонетические символы неполным и неудовлетворительным средством для такого обучения. Одномерные фонетические символы, напечатанные на бумаге, сообщают нашим глазам, какие звуки они представляют, сообщают нашим ушам, какие звуки мы должны произнести, но мало или совсем не обращаются к нашему воображению или даже к нашим телам (кроме наших артикуляторов). Стремясь перенести фонетику в тот же физический мир, что и другие классы перформанса, я много лет работал со студентами-актёрами над тем, как заставить символы прыгать со страницы, входить в наше тело и требовать от нас их выражения. Величайшие драматические рассказчики, от Уильяма Шекспира до Кэрил Черчилль, требуют от нас не только хорошего языкового слуха; они пишут для сенсориума и из него; все тело нервов, которое стимулирует чувственную реакцию. Их тексты бросают вызов актерам и зрителям, вибрируя идеями, конфликтами и страстями с ног до головы. Курс «Фонетическая подушка» направлен на то, чтобы научить актера быть сверхвербальным: одновременно воспринимать язык тактильно, аудиально и образно; действительно, чтобы насладиться вкусами и запахами языка. 908:25 Источником вдохновения для «Фонетических подушек» послужила работа Кристин Линклейтер «Звук и движение», в которой прослеживается язык от самого примитивного импульса до возвышенных текстов Шекспира. Используя различные игры, некоторые из которых заимствованы у Ллинклейтера, я знакомлю учащихся с Международным фонетическим алфавитом в виде подушек в форме символов.
Стремясь перенести фонетику в тот же физический мир, что и другие классы перформанса, я много лет работал со студентами-актёрами над тем, как заставить символы прыгать со страницы, входить в наше тело и требовать от нас их выражения. Величайшие драматические рассказчики, от Уильяма Шекспира до Кэрил Черчилль, требуют от нас не только хорошего языкового слуха; они пишут для сенсориума и из него; все тело нервов, которое стимулирует чувственную реакцию. Их тексты бросают вызов актерам и зрителям, вибрируя идеями, конфликтами и страстями с ног до головы. Курс «Фонетическая подушка» направлен на то, чтобы научить актера быть сверхвербальным: одновременно воспринимать язык тактильно, аудиально и образно; действительно, чтобы насладиться вкусами и запахами языка. 908:25 Источником вдохновения для «Фонетических подушек» послужила работа Кристин Линклейтер «Звук и движение», в которой прослеживается язык от самого примитивного импульса до возвышенных текстов Шекспира. Используя различные игры, некоторые из которых заимствованы у Ллинклейтера, я знакомлю учащихся с Международным фонетическим алфавитом в виде подушек в форме символов. Предварительно я предлагаю, чтобы каждая подушка действительно вибрировала со звуком, который она представляет, и, кроме того, чтобы эти звуки обладали способностью двигать тела учеников. С помощью этого начального воображаемого прыжка учащиеся видят, осязают и придают выразительный голос и физический жест каждой фонеме. Работа одновременно эмоциональная и точная, аналитическая и основанная на действиях. Я не прошу студентов моделировать формы символов своими телами, а скорее позволить своим телам двигаться под действием самого звука. Можно сказать, абстрактные звуки в их телах. Но они не просто разносят звук по комнате; они используют звук как топливо для удовлетворения желания общаться.
Предварительно я предлагаю, чтобы каждая подушка действительно вибрировала со звуком, который она представляет, и, кроме того, чтобы эти звуки обладали способностью двигать тела учеников. С помощью этого начального воображаемого прыжка учащиеся видят, осязают и придают выразительный голос и физический жест каждой фонеме. Работа одновременно эмоциональная и точная, аналитическая и основанная на действиях. Я не прошу студентов моделировать формы символов своими телами, а скорее позволить своим телам двигаться под действием самого звука. Можно сказать, абстрактные звуки в их телах. Но они не просто разносят звук по комнате; они используют звук как топливо для удовлетворения желания общаться.
В начале работы студия оживает какофонией необработанных звуков, когда ученики случайным образом сталкиваются с фонетическими подушками и передают звуки друг другу. Но из хаоса приходит порядок, поскольку упражнения с подушкой продолжаются, учащиеся оттачивают артикуляцию с глубоким интересом к точному формированию и уникальным характеристикам каждой фонемы. Слух тренируется, но никогда не исключаются другие чувства. На самом деле, я часто учу студентов чувствовать звук, а не слышать его. Поскольку фонетика — это актерская работа, она обязательно игровая. Процесс, часто веселый, также чрезвычайно требователен к голосу и телу. Это трудное, волнующее приключение, в котором учащийся заново переживает язык с жадным любопытством младенца. Это новое открытие бесчисленных звуков, которые в начале жизни дергались, тряслись, прокатывались и извивались в теле. Класс Pillow также отвечает обычным требованиям курса фонетики, таким как идентификация звука / символа и фонетическая транскрипция. Существуют специальные игры с подушками для изучения и репетиции текстов, а также для анализа и усвоения диалектов. Много времени тратится на то, чтобы чередовать физические переживания с подушками и записывать то, что помнит тело. На типичном занятии учащиеся используют упражнения с подушками, чтобы разогреть связь между голосом и телом, подробно расшифровывают свои собственные модели речи и речи других людей, тренируются читать свои транскрипции вслух, а затем с новым любопытством погружаются обратно в к интуитивным фонетическим играм с подушками.
Слух тренируется, но никогда не исключаются другие чувства. На самом деле, я часто учу студентов чувствовать звук, а не слышать его. Поскольку фонетика — это актерская работа, она обязательно игровая. Процесс, часто веселый, также чрезвычайно требователен к голосу и телу. Это трудное, волнующее приключение, в котором учащийся заново переживает язык с жадным любопытством младенца. Это новое открытие бесчисленных звуков, которые в начале жизни дергались, тряслись, прокатывались и извивались в теле. Класс Pillow также отвечает обычным требованиям курса фонетики, таким как идентификация звука / символа и фонетическая транскрипция. Существуют специальные игры с подушками для изучения и репетиции текстов, а также для анализа и усвоения диалектов. Много времени тратится на то, чтобы чередовать физические переживания с подушками и записывать то, что помнит тело. На типичном занятии учащиеся используют упражнения с подушками, чтобы разогреть связь между голосом и телом, подробно расшифровывают свои собственные модели речи и речи других людей, тренируются читать свои транскрипции вслух, а затем с новым любопытством погружаются обратно в к интуитивным фонетическим играм с подушками. Таким образом, они отрабатывают технические навыки в игровой, творческой среде актерской студии. К концу курса студент готов претендовать на владение языковыми моделями, требуемыми драматургом, персонажем или диалектом.
Таким образом, они отрабатывают технические навыки в игровой, творческой среде актерской студии. К концу курса студент готов претендовать на владение языковыми моделями, требуемыми драматургом, персонажем или диалектом.
Помимо обучения студентов актерскому мастерству, я часто провожу семинары по фонетической подушке для учителей. В число участников входят преподаватели голоса и речи из колледжей и университетов, а иногда и специалисты в смежных областях, таких как логопедия, речевая патология и среднее образование. Десятки профессиональных обучающих программ владеют набором фонетических подушек. Книги «Радость фонетики и акцентов», в которых излагается подход, и «Оживление речи», сопровождающая рабочая тетрадь, были приняты многими учителями на протяжении многих лет. Учителя сообщают о многих положительных результатах для своих учеников, в том числе об ускорении сроков изучения фонетических символов, увеличении запоминаемости символов, усилении связи с языком и его жизнеспособности, а также о большем энтузиазме в отношении предмета. Фонетические подушки изготавливаются вручную и должны быть либо заказаны на заказ, либо изготовлены в домашних условиях. Многие школы убедили свои магазины костюмов сделать эту работу.
Фонетические подушки изготавливаются вручную и должны быть либо заказаны на заказ, либо изготовлены в домашних условиях. Многие школы убедили свои магазины костюмов сделать эту работу.
Мелани Т. Кушон, доктор философии
Сиссе, Усман Х; Ма, Лян; Деккер, Джон П.; Хиль, Павел П; Юн, Юнг-Хо; Бренчли, Джейсон М; Блэр, Роберт; Пахар, Бапи; Шабе, Магали; Ван Ромпей, Коэн К.А.; Кислер, Ребекка; Сукура, Антти; Хирш, Ванесса; Катти, Гита; Лю, Юэцинь; Пэн, Ли; Чен, Цзе; Песня, июнь; Вайсенбахер-Ланг, Кристиана; Сюй, Цзе; Апхэм, Натан С.; Стаджич, Джейсон Э.; Куомо, Кристина А; Подушка, Мелани Т.; Ковач, Джозеф А. 2021. Геномный взгляд на специфичную для хозяина адаптацию рода Pneumocystis. Биология коммуникации , 4 1, 305
Сиссе, Усман Х; Ма, Лян; Деккер, Джон П.; Хиль, Павел П; Юн, Юнг-Хо; Бренчли, Джейсон М; Блэр, Роберт; Пахар, Бапи; Шабе, Магали; Ван Ромпей, Коэн К.А.; Кислер, Ребекка; Сукура, Антти; Хирш, Ванесса; Катти, Гита; Лю, Юэцинь; Пэн, Ли; Чен, Цзе; Песня, июнь; Вайсенбахер-Ланг, Кристиана; Сюй, Цзе; Апхэм, Натан С. ; Стаджич, Джейсон Э.; Куомо, Кристина А; Подушка, Мелани Т.; Ковач, Джозеф А. 2021. Геномный взгляд на специфичную для хозяина адаптацию рода Pneumocystis. Биология коммуникаций , 4 1, 305
; Стаджич, Джейсон Э.; Куомо, Кристина А; Подушка, Мелани Т.; Ковач, Джозеф А. 2021. Геномный взгляд на специфичную для хозяина адаптацию рода Pneumocystis. Биология коммуникаций , 4 1, 305
Мизель, Линн; Подушка, Мелани Т.; Эшбо, Алан; Лопес, Сантьяго Р.; Онг, Вун, 2021. Эффективность резафунгина в профилактических мышиных моделях инвазивного кандидоза, аспергиллеза и пневмоцистной пневмонии. Противомикробные препараты и химиотерапия , 65 3,
Cushion, Melanie T; Кумар, Параг; Лу, Жуин; Эшбо, Алан; Адеохо, Лилиан В.; Альфаро, Рауль; Маннино, Рафаэль; Трамонт, Эдмунд; Ковач, Джозеф А. 2020. Новый инкохлеированный состав улучшает активность атоваквона в мышиной модели пневмоцистной пневмонии. Журнал инфекционных болезней , ,
Ма, Лян; Чен, Зехуа; Хуан, Да Вэй; Сиссе, Усман Х; Ротенбургер, Джейми Л.; Латинн, Алиса; Бишоп, Лиза; Блэр, Роберт; Бренчли, Джейсон М; Шабе, Магали; Дэн, Силун; Хирш, Ванесса; Кислер, Ребекка; Катти, Гита; Лю, Юэцинь; Марголис, Даниэль; Моран, Серж; Пахар, Бапи; Пэн, Ли; Ван Ромпей, Коэн К.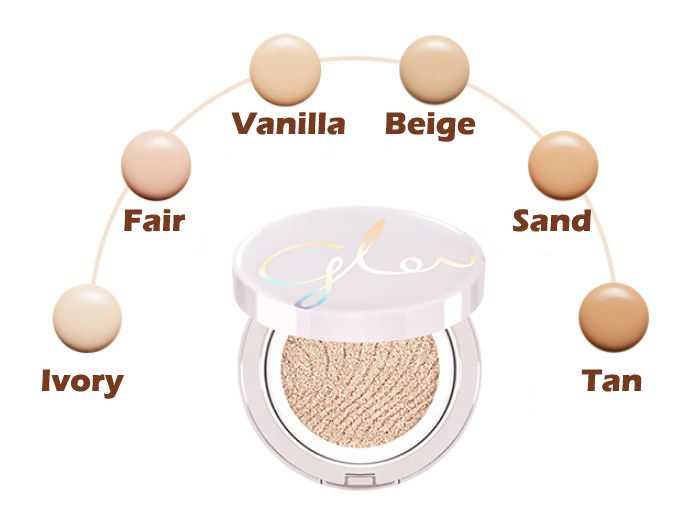 А.; Песня, Сяохун; Песня, июнь; Сукура, Антти; Тапар, Сабрина; Ван, Хунхуэй; Вайсенбахер-Ланг, Кристиана; Сюй, Цзе; Ли, Чао-Хун; Джардин, Клэр; Лемпицки, Ричард А; Подушка, Мелани Т.; Куомо, Кристина А; Ковач, Джозеф А. 2020. Разнообразие и сложность семейства белков большой поверхности в компактных геномах нескольких видов Pneumocystis. mBio , 11 2,
А.; Песня, Сяохун; Песня, июнь; Сукура, Антти; Тапар, Сабрина; Ван, Хунхуэй; Вайсенбахер-Ланг, Кристиана; Сюй, Цзе; Ли, Чао-Хун; Джардин, Клэр; Лемпицки, Ричард А; Подушка, Мелани Т.; Куомо, Кристина А; Ковач, Джозеф А. 2020. Разнообразие и сложность семейства белков большой поверхности в компактных геномах нескольких видов Pneumocystis. mBio , 11 2,
Лю, Гуан-Шэн; Балвег, Ричард; Эшбо, Алан; Чжан, Инь; Фаччоло, Джозеф; Подушка, Мелани Т.; Zhang, Tongli 2019. Исправление к: Модель количественной системной фармакологии (QSP) для лечения Pneumocystis у мышей. BMC системная биология , 13 1, 40
Luraschi, A; Ричард, С; Алмейда, J M G C F; Паньи, М; Подушка, М Т; Hauser, PM 2019. Анализы экспрессии и иммуноокрашивания предполагают, что первичный гомоталлиз пневмоцистиса затрагивает трофические клетки, демонстрирующие как плюсовые, так и минусовые феромонные рецепторы. mBio , 10 4,
Cushion, Melanie T 2018. Ответ Невезу и др., «Увлекательная модель Pneumocystis murina на мышах, обработанных эхинокандином, для понимания Pneumocystis jirovecii». Противомикробные препараты и химиотерапия , 62 8,
Противомикробные препараты и химиотерапия , 62 8,
Cushion, Melanie T; Эшбо, Алан; Хендрикс, Кили; Линке, Майкл Дж.; Тисдейл, Никея; Сэйсон, Стивен Г.; Поролло, Алексей 2018. Экспрессия генов Pneumocystis murina после лечения анидулафунгином приводит к сильным сигналам для полового размножения, целостности клеточной стенки и остановке клеточного цикла, что указывает на необходимость образования асков для пролиферации. Противомикробные препараты и химиотерапия , 62 5,
Cushion, Melanie T; Лимпер, Эндрю Х; Поролло, Алексей; Сапер, Вивиан Э.; Синай, Энтони П.; Вайс, Луи М. 2018. 14-й Международный семинар оппортунистических протистов (IWOP 14). Журнал эукариотической микробиологии , 65 6, 934-939
Gomes, Ana; Феррас, Рикардо; Фикер, Лорен; Коллинз, Маргарет С; Пруденсио, Кристина; Подушка, Мелани Т.; Тейшейра, Катя; Гомес, Паула, 2018 г. Аналоги хлорохина как средства против возбудителей пневмоцистной пневмонии. Противомикробные средства и химиотерапия , 62 11,
Hauser, Philippe M; Кушон, Мелани Т. 2018. Необходим ли секс для распространения и передачи Pneumocystis? Патогены PLoS , 14 12, e1007409
2018. Необходим ли секс для распространения и передачи Pneumocystis? Патогены PLoS , 14 12, e1007409
Lazzarini, Cristina; Харанахалли, Крупананда; Ригер, Роберт; Анантул, Хари Кришна; Десаи, Панкадж Б.; Эшбо, Алан; Линке, Майкл Дж.; Подушка, Мелани Т.; Ружицка, Бела; Хейли, Джон; Одзима, Ивао; Дель Поэта, Маурицио 2018. Ацилгидразоны как противогрибковые агенты, направленные на синтез сфинголипидов грибов. Противомикробные препараты и химиотерапия , 62 5,
Liu, Guan-Sheng; Балвег, Ричард; Эшбо, Алан; Чжан, Инь; Фаччоло, Джозеф; Подушка, Мелани Т.; Zhang, Tongli 2018. Модель количественной системной фармакологии (QSP) для лечения Pneumocystis у мышей. BMC системная биология , 12 1, 77
Мор, Визесато; Релла, Антонелла; Фарно, Амир М.; Сингх, Ашутош; Мунши, Манса; Брайан, Ариэль; Насим, Шамун; Конопка, Джеймс Б.; Одзима, Ивао; Буллесбах, Эрика; Эшбо, Алан; Линке, Майкл Дж.; Подушка, Мелани; Коллинз, Маргарет; Анантул, Хари Кришна; Салланс, Ларри; Десаи, Панкадж Б. ; Видерхольд, Натан П.; Фотергилл, Аннет В.; Киркпатрик, Уильям Р.; Паттерсон, Томас; Вонг, Лай Хун; Синха, Сунита; Гиавер, Гури; Нислоу, Кори; Флаэрти, Патрик; Пан, Сюэвен; Сезар, Габриэле Варгас; де Мело Таварес, Патрисия; Фрейз, Сюзанна; Миранда, Килдэр; Родригес, Марсио Л.; Луберто, Кьяра; Нимрихтер, Леонардо; Дель Поэта, Маурицио, 2018. Опечатка для Mor et al., «Идентификация нового класса противогрибковых препаратов, нацеленных на синтез грибковых сфинголипидов». mBio , 9 2,
; Видерхольд, Натан П.; Фотергилл, Аннет В.; Киркпатрик, Уильям Р.; Паттерсон, Томас; Вонг, Лай Хун; Синха, Сунита; Гиавер, Гури; Нислоу, Кори; Флаэрти, Патрик; Пан, Сюэвен; Сезар, Габриэле Варгас; де Мело Таварес, Патрисия; Фрейз, Сюзанна; Миранда, Килдэр; Родригес, Марсио Л.; Луберто, Кьяра; Нимрихтер, Леонардо; Дель Поэта, Маурицио, 2018. Опечатка для Mor et al., «Идентификация нового класса противогрибковых препаратов, нацеленных на синтез грибковых сфинголипидов». mBio , 9 2,
Мачеевская, Д; Забински, Дж.; Резлер, М; Казмерчак, П; Коллинз, М.С.; Фикер, Л; Cushion, MT 2017. Исправление: разработка высокоактивных бисбензамидинов против Pneumocystis: понимание влияния выбранных заместителей на активность in vitro. МедХимКомм , 8 11, 2164
Мачеевская, Д; Забински, Дж.; Резлер, М; Казмерчак, П; Коллинз, М.С.; Фикер, Л; Cushion, MT 2017. Разработка высокоактивных бисбензамидинов против Pneumocystis: понимание влияния выбранных заместителей на активность in vitro. MedChemComm , 8 10, 2003-2011
MedChemComm , 8 10, 2003-2011
Baker, Frazier N; Подушка, Мелани Т.; Поролло, Алексей 2016. Количественная модель для оценки лекарственной устойчивости патогенов. Journal of fungi (Базель, Швейцария) , 2 4,
Cushion, Melanie T; Коллинз, Маргарет С; Сестерхенн, Томас; Поролло, Алексей; Вадукут, Аниш Кижаккеккара; Мерино, Эдвард Дж. 2016. Функциональная характеристика транспортера инозитола Pneumocystis carinii 1. mBio , 7 6,
Koselny, Kristy; Грин, Джулианна; Дидон, Луи; Холтерман, Джастин П.; Фотергилл, Аннет В.; Видерхольд, Натан П.; Паттерсон, Томас Ф; Подушка, Мелани Т.; Раппель, Чад; Веллингтон, Мелани; Крисан, Дамиан Дж. 2016. Производное целекоксиба AR-12 обладает противогрибковой активностью широкого спектра действия in vitro и улучшает активность флуконазола в мышиной модели криптококкоза. Противомикробные препараты и химиотерапия , 60 12, 7115-7127
Calderon, Enrique J; Подушка, Мелани Т.; Сяо, Лихуа; Лоренцо-Моралес, Джейкоб; Матос, Ольга; Канеширо, Эдна С; Вайс, Луи М. 2015. 13-й Международный семинар оппортунистических протистов (IWOP13). Журнал эукариотической микробиологии , 62 5, 701-9
2015. 13-й Международный семинар оппортунистических протистов (IWOP13). Журнал эукариотической микробиологии , 62 5, 701-9
Mor, Visesato; Релла, Антонелла; Фарно, Амир М.; Сингх, Ашутош; Мунши, Манса; Брайан, Ариэль; Насим, Шамун; Конопка, Джеймс Б.; Одзима, Ивао; Буллесбах, Эрика; Эшбо, Алан; Линке, Майкл Дж.; Подушка, Мелани; Коллинз, Маргарет; Анантул, Хари Кришна; Салланс, Ларри; Десаи, Панкадж Б.; Видерхольд, Натан П.; Фотергилл, Аннет В.; Киркпатрик, Уильям Р.; Паттерсон, Томас; Вонг, Лай Хун; Синха, Сунита; Гиавер, Гури; Нислоу, Кори; Флаэрти, Патрик; Пан, Сюэвен; Сезар, Габриэле Варгас; де Мело Таварес, Патрисия; Фрейз, Сюзанна; Миранда, Килдэр; Родригес, Марсио Л.; Луберто, Кьяра; Нимрихтер, Леонардо; Дель Поэта, Маурицио 2015. Идентификация нового класса противогрибковых препаратов, нацеленных на синтез грибковых сфинголипидов. mBio , 6 3, e00647
Поролло Алексей; Сестерхенн, Томас М; Коллинз, Маргарет С; Велге, Джеффри А; Cushion, Melanie T 2014. Сравнительная геномика видов pneumocystis предполагает отсутствие генов синтеза мио-инозитола и зависимость от транспорта и метаболизма инозитола. mBio , 5 6, e01834
Сравнительная геномика видов pneumocystis предполагает отсутствие генов синтеза мио-инозитола и зависимость от транспорта и метаболизма инозитола. mBio , 5 6, e01834
Подушка, Мелани Т; Кили, Скотт П. 2013. Сборка и аннотация Pneumocystis jirovecii из микробиома легких человека. мБио , 4 2, e00224
Линке, Майкл Дж.; Эшбо, Алан; Коллинз, Маргарет С; Линч, Кили; Кушон, Мелани Т. 2013. Характеристика определенного профиля реакции хозяина на Pneumocystis murina asci во время лечения пневмоцистной пневмонии. Инфекция и иммунитет , 81 3, 984-95
Лобо, Мария Луиза; Эстевес, Франциско; де Соуза, Бруно; Кардосо, Фернандо; Подушка, Мелани Т.; Антунес, Франциско; Матос, Ольга 2013. Терапевтический потенциал каспофунгина в сочетании с триметоприм-сульфаметоксазолом при пневмоцистной пневмонии: пилотное исследование на мышах. PloS one , 8 8, e70619
Weiss, Louis M; Подушка, Мелани Т.; Дидье, Элизабет; Сяо, Лихуа; Марчиано-Кабрал, Франсин; Синай, Энтони П. ; Матос, Ольга; Кальдерон, Энрике Дж.; Канеширо, Эдна С. 2013. 12-й Международный семинар по оппортунистическим протистам (IWOP-12). Журнал эукариотической микробиологии , 60 3, 298-308
; Матос, Ольга; Кальдерон, Энрике Дж.; Канеширо, Эдна С. 2013. 12-й Международный семинар по оппортунистическим протистам (IWOP-12). Журнал эукариотической микробиологии , 60 3, 298-308
Maciejewska, Dorota; Забински, Ежи; Камерчак, Павел; Резлер, Матеуш; Красовска-Вибоцкая, Барбара; Коллинз, Маргарет С; Cushion, Melanie T 2012. Аналоги пентамидина как потенциальные химиотерапевтические средства против Pneumocystis. Европейский журнал медицинской химии , 48 , 164-73
Поролло Алексей; Меллер, Ярослав; Джоши, Йогеш; Джайсвал, Викаш; Смулиан, Джордж; Cushion, Melanie T 2012. Анализ современных противогрибковых средств и их целей в геноме Pneumocystis carinii. Текущие цели по наркотикам , 13 12, 1575-85
Sinai, Anthony P; Канеширо, Эдна С; Уорд, Онорина; Вайс, Луи М.; Кушон, Мелани Т. 2012. Состояние исследований оппортунистических инфекций, связанных со СПИДом, и важность поддержки небольших исследовательских сообществ. Эукариотическая клетка , 11 2, 90-7
Подушка, Мелани Т; Коллинз, Маргарет С. 2011. Восприимчивость Pneumocystis к эхинокандинам в суспензионных и биопленочных культурах. Противомикробные препараты и химиотерапия , 55 10, 4513-8
2011. Восприимчивость Pneumocystis к эхинокандинам в суспензионных и биопленочных культурах. Противомикробные препараты и химиотерапия , 55 10, 4513-8
Cushion, Melanie T 2010. Являются представителями грибкового рода pneumocystis (a) комменсалы; б) оппортунисты; (в) патогены; или (d) все вышеперечисленное? PLoS патогены , 6 9,
Cushion, Melanie T; Линке, Майкл Дж.; Эшбо, Алан; Сестерхенн, Том; Коллинз, Маргарет С; Линч, Кили; Брубейкер, Рональд; Уолцер, Питер Д. 2010. Лечение эхинокандином пневмоцистной пневмонии на моделях грызунов истощает кисты, оставляя трофические отягощения, которые не могут передавать инфекцию. PloS one , 5 1, e8524
Подушка, Мелани Т; Стрингер, Джеймс Р. 2010. Скрытность и оппортунизм: альтернативный образ жизни видов грибов рода Pneumocystis. Ежегодный обзор микробиологии , 64, 431-52
Joffrion, Tiffany M; Коллинз, Маргарет С; Сестерхенн, Томас; Cushion, Melanie T 2010. Функциональная характеристика и локализация ланостеролсинтазы Pneumocystis carinii. Эукариотическая клетка , 9 1, 107-15
Эукариотическая клетка , 9 1, 107-15
Джофрион, Тиффани М; Кушон, Мелани Т. 2010. Биосинтез стеролов и поглощение стеролов грибковым патогеном Pneumocystis carinii. FEMS микробиологические письма , 311 1, 1-9
Sesterhenn, Thomas M; Славен, Брэдли Э.; Кили, Скотт П.; Смулиан, Джордж; Ланг, Б. Франц; Кушон, Мелани Т. 2010. Последовательность и структура линейного митохондриального генома Pneumocystis carinii. Молекулярная генетика и геномика: MGG , 283 1, 63-72
Beck, James M; Cushion, Melanie T. 2009. Семинар по Pneumocystis: резюме к 10-летию. Эукариотическая клетка , 8 4, 446-60
Brubaker, R; Рыжая, SA; Стрингер, Дж. Р.; Кили, С.П.; Подушка, М Т 2009. Дезинформация о Pneumocystis. Клиническая и экспериментальная дерматология , 34 7, e426-7
Подушка, Мелани Т; Коллинз, Маргарет С; Линке, Майкл Дж. 2009. Формирование биопленки Pneumocystis spp. Эукариотическая клетка , 8 2, 197-206
Подушка, Мелани Т; Уолцер, Питер Д. 2009. Открытие доклинических препаратов для новых соединений против пневмоцисты. Текущая медицинская химия , 16 20, 2514-30
2009. Открытие доклинических препаратов для новых соединений против пневмоцисты. Текущая медицинская химия , 16 20, 2514-30
Huang, Tien L; Ванден Эйнде, Жан Жак; Майенс, Энни; Коллинз, Маргарет С; Подушка, Мелани Т.; Раттенди, Донна; Лондоно, Индира; Мазумдер, Лакшман; Бакки, Сайрус Дж.; Ярлетт, Найджел, 2009 г.. Синтез и SAR алкандиамидных бисбензамидинов с антитрипаносомной и антипневмоцистной активностью. Биоорганическая и медицинская химия письма , 19 20, 5884-6
Лю, Ю; Ли, Джессика В.; Бринкманн, Хеннер; Подушка, Мелани Т.; Родригес-Эспелета, Найара; Филипп, Эрве; Ланг, Б. Франц, 2009. Филогеномный анализ подтверждает монофилию Taphrinomycotina, включая делящиеся дрожжи Schizosaccharomyces. Молекулярная биология и эволюция , 26 1, 27-34
Вале, Нуно; Пруденсио, Мигель; Маркес, Катарина А; Коллинз, Маргарет С; Гут, Иржи; Ногейра, Фатима; Матос, Джоана; Розенталь, Филип Дж.; Подушка, Мелани Т.; сделать Росарио, Вирджилио Э.; Мота, Мария М; Морейра, Руи; Gomes, Paula 2009. Имидазохины как противомалярийные и противопневмоцистные средства. Журнал медицинской химии , 52 23, 7800-7
Имидазохины как противомалярийные и противопневмоцистные средства. Журнал медицинской химии , 52 23, 7800-7
Mayence, Annie; Пьетка, Орели; Коллинз, Маргарет С; Подушка, Мелани Т.; Теквани, Бабу Л.; Хуанг, Тиен Л.; Vanden Eynde, Jean Jacques 2008. Новые бисбензимидазолы с противолейшманиальной эффективностью. Буквы по биоорганической и медицинской химии , 18 8, 2658-61
Vale, Nuno; Коллинз, Маргарет С; Гут, Иржи; Феррас, Рикардо; Розенталь, Филип Дж.; Подушка, Мелани Т.; Морейра, Руи; Gomes, Paula 2008. Anti-Pneumocystis carinii и антиплазмодиальная активность имидазолидин-4-онов, полученных из примахина. Буквы по биоорганической и медицинской химии , 18 2, 485-8
Подушка, Мелани Т; Смулиан, Джордж; Славен, Брэдли Э.; Сестерхенн, Том; Арнольд, Джонатан; Стабен, Чак; Поролло, Алексей; Адамчак, Рафал; Меллер, Ярек, 2007. Транскриптом Pneumocystis carinii во время молниеносной инфекции: углеводный обмен и концепция совместимого паразита. PloS one , 2 5, e423
PloS one , 2 5, e423
Подушка, M.T.; Смулян, А.Г.; Славен, BE; Сестерхенн, Т.; Арнольд, Дж.; Стабен, К.; Поролло, А.; Адамчак, Р.; Меллер, Дж. 2007. Транскриптом Pneumocystis carinii во время молниеносной инфекции: углеводный обмен и концепция совместимого паразита PLoS ONE , 2, e423
Keely, Scott P; Линке, Майкл Дж.; Подушка, Мелани Т.; Стрингер, Джеймс Р. 2007. Семейство генов MSG Pneumocystis murina и структура локуса, связанная с его транскрипцией. Генетика и биология грибов: FG & B , 44 9, 905-19
Keely, S.P.; Линке, М.Дж.; Подушка, MT; Стрингер, Дж.Р. 2007. Семейство генов MSG Pneumocystis murina и структура локуса, связанного с его транскрипцией Генетика и биология грибов: FG & B , ,
Keely, S.P.; Линке, М.Дж.; Подушка, MT; Стрингер, Дж.Р. 2007. Семейство генов MSG Pneumocystis murina и структура локуса, связанного с его транскрипцией Генетика и биология грибов: FG & B , 44 9, 905
Тачадо, Сувенир D; Чжан, Цзяньминь; Чжу, Цзиньпин; Патель, Наймиш; Подушка, Мелани; Koziel, Henry 2007. Опосредованное Pneumocystis высвобождение IL-8 макрофагами требует совместной экспрессии маннозных рецепторов и TLR2. Журнал биологии лейкоцитов , 81 1, 205-11
Опосредованное Pneumocystis высвобождение IL-8 макрофагами требует совместной экспрессии маннозных рецепторов и TLR2. Журнал биологии лейкоцитов , 81 1, 205-11
Cushion, Melanie T; Smulian, A George 2006. Pneumocystis 2006: краткое изложение исследования, представленного на девятом Международном семинаре по условно-патогенным простейшим. Журнал эукариотической микробиологии , 53 Приложение 1 , S80-4
Подушка, Мелани Т; Уолцер, Питер Д.; Эшбо, Алан; Ребхольц, Сандра; Брубейкер, Рональд; Ванден Эйнде, Жан Жак; Майенс, Энни; Huang, Tien L 2006. Отбор in vitro и эффективность in vivo бисбензамидинов, связанных с пиперазином и алкандиамидом, против пневмоцистной пневмонии у мышей. Противомикробные средства и химиотерапия , 50 7, 2337-43
Cushion,M.T.; Смулян, А.Г. 2006. Pneumocystis 2006: резюме исследования, представленного на девятом Международном семинаре по оппортунистическим протистам The Journal of eukaryotic microbiology , 53 Suppl 1, S80
Cushion, M. T.; Уолцер, PD; Эшбо, А.; Ребхольц, С.; Брубейкер, Р.; Ванден Эйнде, Дж. Дж.; Майенс, А.; Хуанг, Т.Л. 2006. Отбор in vitro и эффективность пиперазин- и алкандиамид-связанных бисбензамидинов in vivo против Pneumocystis pneumonia у мышей Противомикробные агенты и химиотерапия , 50 7, 2337
T.; Уолцер, PD; Эшбо, А.; Ребхольц, С.; Брубейкер, Р.; Ванден Эйнде, Дж. Дж.; Майенс, А.; Хуанг, Т.Л. 2006. Отбор in vitro и эффективность пиперазин- и алкандиамид-связанных бисбензамидинов in vivo против Pneumocystis pneumonia у мышей Противомикробные агенты и химиотерапия , 50 7, 2337
Dei-Cas, Eduardo; Шабе, Магали; Мухлис, Рая; Дюран-Жоли, Изабель; Алиуат, Эль Мухтар; Стрингер, Джеймс Р.; Подушка, Мелани; Ноэль, Кристоф; де Хоог, Г. Сибрен; Гийо, Жак; Viscogliosi, Eric 2006. Pneumocystis oryctolagi sp. nov., некультивируемый грибок, вызывающий пневмонию у кроликов при отъеме: обзор современных знаний и описание нового таксона на генотипической, филогенетической и фенотипической основе. Микробиологические обзоры FEMS , 30 6, 853-71
Icenhour, Crystal R; Арнольд, Джонатан; Медведович, Марио; Кушон, Мелани Т. 2006. Конкурентное сосуществование двух видов Pneumocystis. Инфекция, генетика и эволюция: журнал молекулярной эпидемиологии и эволюционной генетики инфекционных заболеваний , 6 3, 177-86
Icenhour,C. R.; Арнольд, Дж.; Медведович М.; Подушка, М.Т. 2006. Конкурентное сосуществование двух видов Pneumocystis Инфекция, генетика и эволюция: журнал молекулярной эпидемиологии и эволюционной генетики инфекционных заболеваний , 6 3, 177
R.; Арнольд, Дж.; Медведович М.; Подушка, М.Т. 2006. Конкурентное сосуществование двух видов Pneumocystis Инфекция, генетика и эволюция: журнал молекулярной эпидемиологии и эволюционной генетики инфекционных заболеваний , 6 3, 177
Джоффрион, Тиффани М.; Коллинз, Маргарет С; Cushion, Melanie T 2006. Микроаэрофильные условия повышают жизнеспособность и влияют на реакцию Pneumocystis carinii на лекарства in vitro. The Journal of eukaryotic microbiology , 53 Suppl 1, S117-8
Joffrion, T.M.; Коллинз, MS; Подушка, М.Т. 2006. Микроаэрофильные условия повышают жизнеспособность и влияют на реакцию Pneumocystis carinii на лекарства in vitro The Journal of eukaryotic microbiology , 53 Suppl 1, S117
Канеширо, Эдна С.; Кали, Энн; Подушка, Мелани Т.; Линдси, Дэвид С; Марчиано-Кабрал, Франсин; Мид, Ян Р.; Вайс, Луи М. 2006. Международные семинары оппортунистических протистов. The Journal of eukaryotic microbiology , 53 Suppl 1, S1-7
Redhead, S. A.; Подушка, MT; Френкель, Дж. К.; Стрингер, Дж.Р. 2006. Pneumocystis и Trypanosoma cruzi: номенклатура и типизация Журнал эукариотической микробиологии , 53 1, 2
A.; Подушка, MT; Френкель, Дж. К.; Стрингер, Дж.Р. 2006. Pneumocystis и Trypanosoma cruzi: номенклатура и типизация Журнал эукариотической микробиологии , 53 1, 2
Sesterhenn, Thomas M; Подушка, Мелани Т.; Славен, Брэдли Э.; Смулян, А. Джордж, 2006. Последовательность митохондриального генома Pneumocystis carinii: значение для биологической функции и идентификация потенциальных мишеней для лекарств. The Journal of eukaryotic microbiology , 53 Suppl 1, S154-5
Sesterhenn, T.M.; Подушка, MT; Славен, BE; Смулян, А.Г. 2006. Последовательность митохондриального генома Pneumocystis carinii: значение для биологической функции и идентификация потенциальных мишеней для лекарств Журнал эукариотической микробиологии , 53 Дополнение 1, S154
Slaven, Bradley E; Меллер, Ярослав; Поролло, Алексей; Сестерхенн, Томас; Смулиан, Джордж; Кушон, Мелани Т. 2006. Проект сборки и аннотации генома Pneumocystis carinii. The Journal of eukaryotic microbiology , 53 Suppl 1, S89-91
Slaven, Bradley E; Поролло, Алексей; Сестерхенн, Томас; Смулиан, Джордж; Подушка, Мелани Т. ; Меллер, Ярослав 2006. Крупномасштабная характеристика интронов в геноме Pneumocystis carinii. Журнал эукариотической микробиологии , 53 Приложение 1, S151-3
; Меллер, Ярослав 2006. Крупномасштабная характеристика интронов в геноме Pneumocystis carinii. Журнал эукариотической микробиологии , 53 Приложение 1, S151-3
Slaven, B.E.; Меллер, Дж.; Поролло, А.; Сестерхенн, Т.; Смулян, А.Г.; Подушка, М.Т. 2006. Проект сборки и аннотации генома Pneumocystis carinii 9.0003 Журнал эукариотической микробиологии , 53 Приложение 1, S89
Slaven, B.E.; Поролло, А.; Сестерхенн, Т.; Смулян, А.Г.; Подушка, MT; Меллер, Дж. 2006. Крупномасштабная характеристика интронов в геноме Pneumocystis carinii The Journal of eukaryotic microbiology , 53 Suppl 1, S151
Cushion, Melanie T; Стрингер, Джеймс Р. 2005. Действительно ли имя было изменено? Это имеет место для большинства исследователей. Клинические инфекционные заболевания: официальная публикация Американского общества инфекционистов , 41 12, 1756-8
Подушка, M.T.; Стрингер, Дж.Р. 2005. Действительно ли изменилось название? Для большинства исследователей он имеет Клинические инфекционные заболевания: официальная публикация Американского общества инфекционных заболеваний , 41 12, 1756
Keely, Scott P; Рено, Юбер; Уэйкфилд, Энн Э. ; Подушка, Мелани Т.; Смулиан, Джордж; Фоскер, Найджел; Фрейзер, Одри; Харрис, Дэвид; Мерфи, Ли; Прайс, Клэр; Перепел, Майкл А; Сигер, Кэти; Шарп, Сара; Тиндал, Кэролайн Дж.; Уоррен, Тим; Зейдервейк, Эдуард; Баррелл, Барклай Г.; Стрингер, Джеймс Р.; Холл, Нил, 2005. Массивы генов на теломерах Pneumocystis carinii. Genetics , 170 4, 1589-600
; Подушка, Мелани Т.; Смулиан, Джордж; Фоскер, Найджел; Фрейзер, Одри; Харрис, Дэвид; Мерфи, Ли; Прайс, Клэр; Перепел, Майкл А; Сигер, Кэти; Шарп, Сара; Тиндал, Кэролайн Дж.; Уоррен, Тим; Зейдервейк, Эдуард; Баррелл, Барклай Г.; Стрингер, Джеймс Р.; Холл, Нил, 2005. Массивы генов на теломерах Pneumocystis carinii. Genetics , 170 4, 1589-600
Keely, S.P.; Рено, Х .; Уэйкфилд, AE; Подушка, MT; Смулян, А.Г.; Фоскер, Н.; Фрейзер, А.; Харрис, Д.; Мерфи, Л.; Цена, с.; Перепел, Массачусетс; Сигер, К.; Шарп, С.; Тиндал, CJ; Уоррен, Т.; Зуидервейк, Э.; Баррелл, Б.Г.; Стрингер, младший; Холл, Н. 2005. Массивы генов на теломерах Pneumocystis carinii Genetics , 1704, 1589
Peterson, J Chad; Кушон, Мелани Т. 2005. Пневмоцистоз: не только пневмония. Текущее мнение в микробиологии , 8 4, 393-8
Peterson, J.C.; Подушка, М.Т. 2005. Pneumocystis: не только пневмония Современное мнение в области микробиологии , 8 4, 393
Zhang, Jianmin; Тачадо, Сувенир Д; Патель, Наймиш; Чжу, Цзиньпин; Имрих, Эми; Манфруэлли, Паскаль; Подушка, Мелани; Кинан, Т. Бернар; Koziel, Henry 2005. Отрицательная регулирующая роль маннозных рецепторов в высвобождении провоспалительных цитокинов альвеолярными макрофагами человека in vitro. Журнал биологии лейкоцитов , 78 3, 665-74
Бернар; Koziel, Henry 2005. Отрицательная регулирующая роль маннозных рецепторов в высвобождении провоспалительных цитокинов альвеолярными макрофагами человека in vitro. Журнал биологии лейкоцитов , 78 3, 665-74
Чжан, Цзяньминь; Чжу, Цзиньпин; Бу, Ся; Подушка, Мелани; Кинан, Т. Бернар; Авраам, Хава; Koziel, Henry 2005. Активация Cdc42 и RhoB необходима для маннозного рецептор-опосредованного фагоцитоза альвеолярными макрофагами человека. Молекулярная биология клетки , 16 2, 824-34
Кушон, Мелани Т. 2004. Пневмоцистис: распуская покров мрака. Тенденции в микробиологии , 12 5, 243-9
Кушон, Мелани Т; Уолцер, Питер Д.; Коллинз, Маргарет С; Ребхольц, Сандра; Ванден Эйнде, Жан Жак; Майенс, Энни; Huang, Tien L 2004. Высокоактивные соединения против Pneumocystis carinii в библиотеке новых бисбензамидинов, связанных пиперазином, и родственных соединений. Противомикробные препараты и химиотерапия , 48 11, 4209-16
Cushion, M. T. 2004. Pneumocystis: раскрывая покров мрака Тенденции в микробиологии , 12 5, 243
T. 2004. Pneumocystis: раскрывая покров мрака Тенденции в микробиологии , 12 5, 243
Cushion, M.T. 2004. Сравнительная геномика Pneumocystis carinii с другими простейшими: последствия для образа жизни Журнал эукариотической микробиологии , 51 1, 30
Cushion, MT; Уолцер, PD; Коллинз, MS; Ребхольц, С.; Ванден Эйнде, Дж. Дж.; Майенс, А.; Хуанг, Т.Л. 2004. Высокоактивные соединения против Pneumocystis carinii в библиотеке новых бисбензамидинов, связанных пиперазином, и родственных соединений Антимикробные препараты и химиотерапия , 48 11, 4209
Keely, Scott P; Фишер, Джаред М; Подушка, Мелани Т.; Стрингер, Джеймс Р. 2004. Филогенетическая идентификация Pneumocystis murina sp. nov., новый вид лабораторных мышей. Microbiology (Reading, England) , 150 Pt 5, 1153-65
Keely, S.P.; Фишер, Дж. М.; Подушка, MT; Стрингер, Дж.Р. 2004. Филогенетическая идентификация Pneumocystis murina sp. nov., новый вид лабораторных мышей Микробиология (Рединг, Англия) , 150 Pt 5, 1153
Майенс, Энни; Ванден Эйнде, Жан Жак; Крогстад, Фрэн М. ; Крогстад, Дональд Дж.; Подушка, Мелани Т.; Huang, Tien L 2004. Параллельный синтез конформационно ограниченных конгенеров пентамидина в фазе раствора и оценка их антиплазмодиальной активности. Журнал медицинской химии , 47 10, 2700-5
; Крогстад, Дональд Дж.; Подушка, Мелани Т.; Huang, Tien L 2004. Параллельный синтез конформационно ограниченных конгенеров пентамидина в фазе раствора и оценка их антиплазмодиальной активности. Журнал медицинской химии , 47 10, 2700-5
Ванден Эйнде, Жан Жак; Майенс, Энни; Хуанг, Тиен Л.; Коллинз, Маргарет С; Ребхольц, Сандра; Уолцер, Питер Д.; Cushion, Melanie T 2004. Новые бисбензамидины как потенциальные кандидаты в лекарства для лечения пневмонии, вызванной Pneumocystis carinii. письма по биоорганической и медицинской химии , 14 17, 4545-8
Vanden Eynde, J.J.; Майенс, А.; Хуанг, Т.Л.; Коллинз, MS; Ребхольц, С.; Уолцер, PD; Подушка, М.Т. 2004. Новые бисбензамидины как потенциальные кандидаты в лекарства для лечения пневмонии Pneumocystis carinii Письма по биоорганической и медицинской химии , 14 17, 4545
Zhang, Jianmin; Чжу, Цзиньпин; Имрих, Эми; Подушка, Мелани; Кинан, Т. Бернар; Koziel, Henry 2004. Pneumocystis активирует передачу сигналов NF-kappaB альвеолярных макрофагов человека через маннозные рецепторы. Инфекция и иммунитет , 72 6, 3147-60
Инфекция и иммунитет , 72 6, 3147-60
Collins, Margaret S; Бансил, Сандип; Cushion, Melanie T 2003. Профилирование экспрессии реакций Pneumocystis carinii на медикаментозное лечение с использованием ДНК-макрочипов. The Journal of eukaryotic microbiology , 50 Suppl, 605-6
Collins, M.S.; Бансил, С.; Подушка, М.Т. 2003. Профилирование экспрессии реакций Pneumocystis carinii на медикаментозное лечение с использованием ДНК-макрочипов Журнал эукариотической микробиологии , 50 Suppl, 605
Кили, Скотт П.; Подушка, Мелани Т.; Stringer, James R 2003. Разнообразие локуса, связанное с транскрипцией вариабельного поверхностного антигена Pneumocystis carinii, как показатель структуры и динамики популяции инфицированных крыс. Инфекция и иммунитет , 71 1, 47-60
Keely, S.P.; Подушка, MT; Стрингер, Дж.Р. 2003. Разнообразие в локусе, ассоциированном с транскрипцией вариабельного поверхностного антигена Pneumocystis carinii, как показатель структуры и динамики популяции инфицированных крыс Инфекция и иммунитет , 71 1, 47
Linke, Michael J; Ребхольц, Сэнди; Коллинз, Маргарет; Танака, Рейко; Кушон, Мелани Т. 2003. Неинвазивный метод мониторинга пневмонии, вызванной Pneumocystis carinii. Возникающие инфекционные заболевания , 9 12, 1613-6
2003. Неинвазивный метод мониторинга пневмонии, вызванной Pneumocystis carinii. Возникающие инфекционные заболевания , 9 12, 1613-6
Linke,M.J.; Ребхольц, С.; Коллинз, М.; Танака Р.; Подушка, М.Т. 2003. Неинвазивный метод мониторинга пневмоцистной пневмонии Новые инфекционные заболевания , 9 12, 1613
Sesterhenn, Thomas M; Славен, Брэдли Э.; Смулиан, Джордж; Кушон, Мелани Т. 2003. Создание библиотек секвенирования для проекта Pneumocystis Genome. The Journal of eukaryotic microbiology , 50 Suppl, 663-5
Sesterhenn, T.M.; Славен, BE; Смулян, А.Г.; Подушка, М.Т. 2003. Создание библиотек секвенирования для проекта Pneumocystis Genome The Journal of eukaryotic microbiology , 50 Suppl, 663
Xu, Zheng; Лэнс, Бриттон; Варгас, Клаудия; Арпинар, Будак; Бхандаркар, Сучендра; Кремер, Эйлин; Кочут, Крыс Дж.; Миллер, Джон А; Вагнер, Джефф Р.; Вайз, Майкл Дж.; Вундерлих, Джон К.; Стрингер, Джеймс; Смулиан, Джордж; Подушка, Мелани Т. ; Арнольд, Джонатан, 2003. Картирование путем секвенирования генома Pneumocystis с использованием инструмента упорядочения последовательностей ДНК V3. Генетика , 163 4, 1299-313
; Арнольд, Джонатан, 2003. Картирование путем секвенирования генома Pneumocystis с использованием инструмента упорядочения последовательностей ДНК V3. Генетика , 163 4, 1299-313
Xu,Z.; Лэнс, Б.; Варгас, К.; Арпинар, Б.; Бхандаркар С.; Кремер, Э.; Кочут, К.Дж.; Миллер, Дж. А.; Вагнер, младший; Вайс, М.Дж.; Вундерлих, Дж. К.; Стрингер, Дж.; Смулиан, Г.; Подушка, MT; Арнольд, Дж. 2003. Картирование путем секвенирования генома Pneumocystis с использованием инструмента упорядочения последовательностей ДНК V3 Genetics , 1634, 1299
Cornillot, Emmanuel; Келлер, Брайан; Подушка, Мелани Т.; Метенье, Ги; Виварес, Кристиан П. 2002. Точный анализ Pneumocystis carinii f. сп. carinii методом двумерного гель-электрофореза в импульсном поле. Gene , 293 1-2, 87-95
Cornillot, E.; Келлер, Б.; Подушка, MT; Метенье, Г.; Виварес, К.П. 2002. Точный анализ Pneumocystis carinii f. сп. carinii с помощью двумерного гель-электрофореза в пульсирующем поле Gene , 293 1-2, 87
De Lucca, A J; Бланд, Дж. М.; Виго, CB; Подушка, М; Селитренников, С.П.; Питер, Дж; Walsh, TJ 2002. CAY-I, фунгицидный сапонин из Capsicum sp. фрукты. Медицинская микология , 40 2, 131-7
М.; Виго, CB; Подушка, М; Селитренников, С.П.; Питер, Дж; Walsh, TJ 2002. CAY-I, фунгицидный сапонин из Capsicum sp. фрукты. Медицинская микология , 40 2, 131-7
Ледяной час, Кристалл R; Ребхольц, Сандра Л.; Коллинз, Маргарет С; Cushion, Melanie T 2002. Раннее заражение Pneumocystis carinii у новорожденных крыс, о чем свидетельствует ПЦР и мазки из полости рта. Эукариотическая клетка , 1 3, 414-9
Icenhour, C.R.; Ребхольц, С.Л.; Коллинз, MS; Подушка, М.Т. 2002. Раннее заражение Pneumocystis carinii у новорожденных крыс по данным ПЦР и мазков из полости рта Эукариотическая клетка , 1 3, 414
Zhou, Wenxu; Нгуен, Тхи Туи Минь; Коллинз, Маргарет С; Подушка, Мелани Т.; Nes, W David 2002. Доказательства наличия множественных путей стеролметилтрансферазы у Pneumocystis carinii. Липиды , 37 12, 1177-86
Zhou,W.; Нгуен, Т.Т.; Коллинз, MS; Подушка, MT; Нес, В.Д. 2002. Доказательства множественных путей стеролметилтрансферазы у Pneumocystis carinii 9. 0003 Липиды , 37 12, 1177
0003 Липиды , 37 12, 1177
Collins, M S; Cushion, MT 2001. Стандартизация анализа скрининга лекарств in vitro с использованием криоконсервированных и охарактеризованных популяций Pneumocystis carinii. The Journal of eukaryotic microbiology , Suppl, 178S-179S
Collins, M.S.; Подушка, М.Т. 2001. Стандартизация скрининга лекарств in vitro с использованием криоконсервированных и охарактеризованных популяций Pneumocystis carinii The Journal of eukaryotic microbiology , Suppl, 178S
Подушка, M T; Бек, Дж. М. 2001. Резюме исследования Pneumocystis, представленное на 7-м Международном семинаре по условно-патогенным простейшим. Журнал эукариотической микробиологии , Приложение, 101S-105S
Cushion, MT; Орр, С; Кили, С.П.; Stringer, JR 2001. Время между прививками и кариотипом форм Pneumocystis carinii f. сп. Carinii влияют на исход экспериментальных коинфекций у крыс. Инфекция и иммунитет , 69 1, 97-107
Подушка, М Т; Смулиан, А. Г. 2001. Проект генома пневмоцистиса: обновление и проблемы. Журнал эукариотической микробиологии , Приложение, 182S-183S
Г. 2001. Проект генома пневмоцистиса: обновление и проблемы. Журнал эукариотической микробиологии , Приложение, 182S-183S
Cushion, M.T.; Бек, Дж.М. 2001. Резюме исследования Pneumocystis, представленное на 7-м Международном семинаре по условно-патогенным простейшим The Journal of eukaryotic microbiology , Suppl, 101S
Cushion, MT; Орр, С.; Кили, С.П.; Стрингер, Дж.Р. 2001. Время между прививками и кариотип формы Pneumocystis carinii f. сп. Carinii влияют на исход экспериментальных коинфекций у крыс Инфекция и иммунитет , 69 1, 97
Подушка, M.T.; Смулян, А.Г. 2001. Проект генома пневмоцистиса: обновление и выпуски Журнал эукариотической микробиологии , Дополнение, 182S
Icenhour, CR; Cushion, MT 2001. Предполагаемая трансмиссивная форма Pneumocystis carinii f. сп. карины. The Journal of eukaryotic microbiology , Suppl, 139S-140S
Icenhour, CR; Ребхольц, С.Л.; Коллинз, М.С.; Cushion, MT 2001. Широкое распространение Pneumocystis carinii в колониях коммерческих крыс, обнаруженное с помощью целенаправленной ПЦР и мазков из полости рта. Журнал клинической микробиологии , 39 10, 3437-41
Журнал клинической микробиологии , 39 10, 3437-41
Icenhour, CR; Ребхольц, С.Л.; Коллинз, М.С.; Cushion, MT 2001. Раннее обнаружение Pneumocystis carinii у новорожденных крыс с помощью целевой ПЦР и оральных мазков. The Journal of eukaryotic microbiology , Suppl, 135S-136S
Icenhour,C.R.; Подушка, М.Т. 2001. Предполагаемая трансмиссивная форма Pneumocystis carinii f. сп. carinii The Journal of eukaryotic microbiology , Suppl, 139S
Icenhour,C.R.; Ребхольц, С.Л.; Коллинз, MS; Подушка, М.Т. 2001 г. Раннее выявление Pneumocystis carinii у новорожденных крыс с помощью направленной ПЦР и мазков из ротовой полости The Journal of eukaryotic microbiology , Suppl, 135S
Icenhour, C.R.; Ребхольц, С.Л.; Коллинз, MS; Подушка, М.Т. 2001. Широкое распространение Pneumocystis carinii в коммерческих колониях крыс, обнаруженное с помощью целевой ПЦР и мазков из полости рта Journal of Clinical Microbiology , 39 10, 3437
Keely, S P; Уэйкфилд, AE; Подушка, М Т; Смулян, А. Г.; Холл, Н.; Баррелл, Б. Г.; Стрингер, Дж. Р. 2001. Подробная структура концов хромосом Pneumocystis carinii. Журнал эукариотической микробиологии , Приложение, 118S-120S
Г.; Холл, Н.; Баррелл, Б. Г.; Стрингер, Дж. Р. 2001. Подробная структура концов хромосом Pneumocystis carinii. Журнал эукариотической микробиологии , Приложение, 118S-120S
Кили, С.П.; Уэйкфилд, AE; Подушка, MT; Смулян, А.Г.; Холл, Н.; Баррелл, Б.Г.; Стрингер, Дж.Р. 2001. Подробная структура концов хромосом Pneumocystis carinii The Journal of eukaryotic microbiology , Suppl, 118S
Nahimana, A; Подушка, М Т; Блан, D S; Hauser, PM 2001. Быстрый метод полиморфизма конформации одноцепочечной ПЦР для дифференциации и оценки относительной численности особых форм Pneumocystis carinii, инфицирующих крыс. Журнал клинической микробиологии , 39 12, 4563-5
Нахимана, А.; Подушка, MT; Блан, Д.С.; Хаузер, П.М. 2001. Быстрый метод полиморфизма конформации одноцепочечной ПЦР для дифференциации и оценки относительной численности особых форм Pneumocystis carinii, заражающих крыс Journal of Clinical Microbiology , 39 12, 4563
Rebholz, S L; Cushion, MT 2001. Три новые формы кариотипа Pneumocystis carinii f. сп. carinii, идентифицированный с помощью электрофореза в однородном электрическом поле с фиксированным контуром (CHEF). The Journal of eukaryotic microbiology , Suppl, 109S-110S
Три новые формы кариотипа Pneumocystis carinii f. сп. carinii, идентифицированный с помощью электрофореза в однородном электрическом поле с фиксированным контуром (CHEF). The Journal of eukaryotic microbiology , Suppl, 109S-110S
Rebholz, S.L.; Подушка, М.Т. 2001. Три новые формы кариотипа Pneumocystis carinii f. сп. carinii, идентифицированный с помощью электрофореза в однородном электрическом поле с фиксированным контуром (CHEF) The Journal of eukaryotic microbiology , Suppl, 109S
Smulian, AG; Сестерхенн, Т; Танака, Р; Cushion, MT 2001. Ген рецептора феромона ste3 Pneumocystis carinii окружен кластером генов передачи сигнала. Генетика , 157 3, 991-1002
Смулян А.Г.; Сестерхенн, Т.; Танака Р.; Подушка, М.Т. 2001. Ген рецептора феромона ste3 Pneumocystis carinii окружен кластером генов трансдукции сигнала Генетика , 1573, 991
Stringer, JR; Подушка, М Т; Wakefield, AE 2001. Новая номенклатура рода Pneumocystis. The Journal of eukaryotic microbiology , Suppl, 184S-189S
Stringer, J. R.; Подушка, MT; Уэйкфилд, А.Э. 2001. Новая номенклатура рода Pneumocystis 9.0003 The Journal of eukaryotic microbiology , Suppl, 184S
R.; Подушка, MT; Уэйкфилд, А.Э. 2001. Новая номенклатура рода Pneumocystis 9.0003 The Journal of eukaryotic microbiology , Suppl, 184S
Walzer, PD; Эшбо, А; Коллинз, М; Cushion, MT 2001. Препараты против вируса иммунодефицита человека неэффективны против Pneumocystis carinii in vitro и in vivo. Журнал инфекционных болезней , 184 10, 1355-7
Walzer, PD; Эшбо, А; Коллинз, М; Cushion, MT 2001. Эффекты хинупристин-далфопристин in vitro и in vivo против Pneumocystis carinii. Антимикробные препараты и химиотерапия , 45 11, 3234-7
Уолцер, П.Д.; Эшбо, А.; Коллинз, М.; Подушка, М.Т. 2001. Препараты против вируса иммунодефицита человека неэффективны против Pneumocystis carinii in vitro и in vivo. Эшбо, А.; Коллинз, М.; Подушка, М.Т. 2001. In vitro и in vivo эффекты хинупристин-дальфопристин против Pneumocystis carinii Антимикробные агенты и химиотерапия , 45 11, 3234
Cushion, MT; Коллинз, М; Хазра, Б; Kaneshiro, ES 2000. Влияние атоваквона и препаратов на основе диоспирина на клеточный АТФ Pneumocystis carinii f. сп. карины. Противомикробные препараты и химиотерапия , 44 3, 713-9
сп. карины. Противомикробные препараты и химиотерапия , 44 3, 713-9
Cushion,M.T.; Коллинз, М.; Хазра, Б.; Канеширо, Э.С. 2000. Влияние атоваквона и препаратов на основе диоспирина на клеточный АТФ Pneumocystis carinii f. сп. carinii Противомикробные препараты и химиотерапия , 44 3, 713
Kaneshiro, E S; Коллинз, М.С.; Cushion, MT 2000. Ингибиторы биосинтеза стеролов и амфотерицин В снижают жизнеспособность pneumocystis carinii f. сп. карины. Антимикробные препараты и химиотерапия , 44 6, 1630-8
Канеширо, Э.С.; Коллинз, MS; Подушка, М.Т. 2000. Ингибиторы биосинтеза стеролов и амфотерицин В снижают жизнеспособность pneumocystis carinii f. сп. carinii Антимикробные препараты и химиотерапия , 44 6, 1630
Cushion,M.T. 1999. Резюме исследования Pneumocystis, представленное на 6-м Международном семинаре по условно-патогенным простейшим Журнал эукариотической микробиологии , 46 5, 85S
Cushion, M.T.; Линке, М. Дж.; Коллинз, М.; Кили, С.П.; Стрингер, Дж.Р. 1999. Минимальное количество Pneumocystis carinii f. сп. carinii, необходимое для установления инфекций, очень мало The Journal of eukaryotic microbiology , 46 5, 111S
Дж.; Коллинз, М.; Кили, С.П.; Стрингер, Дж.Р. 1999. Минимальное количество Pneumocystis carinii f. сп. carinii, необходимое для установления инфекций, очень мало The Journal of eukaryotic microbiology , 46 5, 111S
Icenhour,C.R.; Арнольд, Дж.; Подушка, М.Т. 1999. Взаимодействия двух популяций Pneumocystis carinii в легких крыс Журнал эукариотической микробиологии , 46 5, 107S
Kaneshiro, E.S.; Коллинз, М.; Подушка, М.Т. 1999. Влияние ингибиторов стеролов на содержание АТФ в Pneumocystis carinii 9.0003 Журнал эукариотической микробиологии , 46 5, 142S
Keely, S.P.; Подушка, MT; Стрингер, Дж.Р. 1999. Определение максимальной частоты генетических перестроек, связанных с вариацией поверхностного антигена Pneumocystis carinii The Journal of eukaryotic microbiology , 465, 128S
Palmer, RJ; Подушка, М Т; Wakefield, AE 1999. Дискриминация Pneumocystis carinii f. крысиного происхождения. сп. Carinii и Pneumocystis carinii f. сп. Ратти с помощью полимеразной цепной реакции. Молекулярные и клеточные зонды , 13 2, 147-55
Молекулярные и клеточные зонды , 13 2, 147-55
Palmer, R.J.; Подушка, MT; Уэйкфилд, А.Э. 1999. Дискриминация Pneumocystis carinii f. крысиного происхождения. сп. Carinii и Pneumocystis carinii f. сп. Ratti с использованием полимеразной цепной реакции Молекулярные и клеточные зонды , 13 2, 147
Weisbroth, SH; Гейстфельд, Дж.; Вайсброт, С.П.; Уильямс, Б; Фельдман, С.Х.; Линке, М.Дж.; Орр, С; Cushion, MT 1999. Латентная инфекция Pneumocystis carinii в коммерческих колониях крыс: сравнение индуктивных иммунодепрессантов плюс гистопатология, ПЦР и серология в качестве методов обнаружения. Журнал клинической микробиологии , 37 5, 1441-6
Weisbroth, S.H.; Гейстфельд, Дж.; Вайсброт, С.П.; Уильямс, Б.; Фельдман, С.Х.; Линке, М.Дж.; Орр, С.; Подушка, М.Т. 1999. Латентная инфекция Pneumocystis carinii в коммерческих колониях крыс: сравнение индуктивных иммунодепрессантов плюс гистопатология, ПЦР и серология в качестве методов обнаружения Журнал клинической микробиологии , 37 5, 1441
Cushion, MT 1998. Таксономия, генетическая организация и жизнь цикл Pneumocystis carinii. Семинары по респираторным инфекциям , 13 4, 304-12
Таксономия, генетическая организация и жизнь цикл Pneumocystis carinii. Семинары по респираторным инфекциям , 13 4, 304-12
Cushion, MT 1998. Генетическая гетерогенность Pneumocystis крысиного происхождения. FEMS Иммунология и медицинская микробиология , 22 1-2, 51-8
Cushion, M.T. 1998. Таксономия, генетическая организация и жизненный цикл Pneumocystis carinii Семинары по респираторным инфекциям , 13 4, 304
Cushion, M.T. 1998. Генетическая гетерогенность Pneumocystis крысиного происхождения Иммунология и медицинская микробиология , 22 1-2, 51
Стрингер, Дж. Р.; Cushion, MT 1998. Геном Pneumocystis carinii. FEMS Иммунология и медицинская микробиология , 22 1-2, 15-26
Stringer, J.R.; Подушка, М.Т. 1998. Геном Pneumocystis carinii FEMS иммунология и медицинская микробиология , 22 1-2, 15
Arnold, J.; Подушка, М.Т. 1997. Создание физической карты генома Pneumocystis Журнал эукариотической микробиологии , 446, 8S
Beach, DH; Чен, Ф; Подушка, М Т; Макомбер, Р. С.; Круди, Г.А.; Видер, Массачусетс; Канеширо, E S 1997. Влияние производных стероидной алленовой фосфоновой кислоты на паразитических протистов Leishmania donovani, Leishmania mexicana mexicana и Pneumocystis carinii carinii. Противомикробные средства и химиотерапия , 41 1, 162-8
С.; Круди, Г.А.; Видер, Массачусетс; Канеширо, E S 1997. Влияние производных стероидной алленовой фосфоновой кислоты на паразитических протистов Leishmania donovani, Leishmania mexicana mexicana и Pneumocystis carinii carinii. Противомикробные средства и химиотерапия , 41 1, 162-8
Beach, D.H.; Чен, Ф.; Подушка, MT; Макомбер, Р.С.; Круди, Г.А.; Видер, Массачусетс; Канеширо, Э.С. 1997. Влияние производных стероидной алленовой фосфоновой кислоты на паразитических протистов Leishmania donovani, Leishmania mexicana mexicana и Pneumocystis carinii carinii 9.0003 Противомикробные средства и химиотерапия , 41 1, 162
Подушка, M T; Чен, Ф; Kloepfer, N 1997. Анализ цитотоксичности для оценки агентов-кандидатов против Pneumocystis carinii. Противомикробные средства и химиотерапия , 41 2, 379-84
Cushion, M T; Уолцер, П. Д.; Смулян, А. Г.; Линке, М.Дж.; Руффоло, Дж. Дж.; Канеширо, Э.С.; Стрингер, Дж. Р. 1997. Терминология жизненного цикла Pneumocystis carinii. Инфекция и иммунитет , 65 10, 4365; автор ответ 4
Инфекция и иммунитет , 65 10, 4365; автор ответ 4
Подушка, M.T.; Арнольд, Дж. 1997. Предложение по проекту генома Pneumocystis The Journal of eukaryotic microbiology , 446, 7S
Cushion, M.T.; Чен, Ф.; Клопфер, Н. 1997. Анализ цитотоксичности для оценки потенциальных агентов против Pneumocystis carinii Antimicrobial Agents and Chemotherapy , 41 2, 379
Cushion, M.T.; Орр, С.; Арнольд, Дж. 1997. Взаимодействия между двумя популяциями Pneumocystis в пределах одного хозяина Журнал эукариотической микробиологии , 44 6, 9S
Подушка, M.T.; Уолцер, PD; Смулян, А.Г.; Линке, М.Дж.; Руффоло, Дж. Дж.; Канеширо, Э.С.; Стрингер, Дж.Р. 1997. Терминология жизненного цикла Pneumocystis carinii Инфекция и иммунитет , 65 10, 4365; ответ автора 4366
Perlada, D E; Смулян, А. Г.; Cushion, MT 1997. Молекулярная эпидемиология и чувствительность энтерококков к антибиотикам в Цинциннати, Огайо: перспективное общегородское исследование. Журнал клинической микробиологии , 35 9, 2342-7
Журнал клинической микробиологии , 35 9, 2342-7
Стрингер-младший; Уэйкфилд, AE; Подушка, MT; Дей-Кас, Э. 1997. Таксономия и номенклатура Pneumocystis: обновление Журнал эукариотической микробиологии , 44 6, 5S
Cushion, MT; Орр, С. 1996. Кинетика 2 генетически различных популяций Pneumocystis carinii в колониях крыс Журнал эукариотической микробиологии , 435, 46S
Cushion, M.T.; Уолцер, П.Д. 1996. Заключительные комментарии: ретроспективная перспектива 4-й Международный семинар по Pneumocystis Журнал эукариотической микробиологии , 43 5, 60S
Keely, S.P.; Подушка, MT; Стрингер, Дж.Р. 1996. Стабильность четырех генетических локусов Pneumocystis carinii sp. ф. carinii Журнал эукариотической микробиологии , 43 5, 49S
Смулян А.Г.; Райан, М; Стабен, К; Cushion, M 1996. Передача сигнала у Pneumocystis carinii: характеристика генов (pcg1), кодирующих альфа-субъединицу G-белка (PCG1) Pneumocystis carinii carinii и Pneumocystis carinii ratti. Инфекция и иммунитет , 64 3, 691-701
Инфекция и иммунитет , 64 3, 691-701
Vasquez, J; Смулян, А. Г.; Линке, М.Дж.; Cushion, MT 1996. Антигенные различия, связанные с генетически отличной Pneumocystis carinii от крыс. Инфекция и иммунитет , 64 1, 290-7
Vasquez, J.; Смулян, А.Г.; Линке, М.Дж.; Подушка, М.Т. 1996. Антигенные различия, связанные с генетически отличной Pneumocystis carinii от крыс Инфекция и иммунитет , 64 1, 290
O’Leary, T J; Цай, М М; Райт, CF; Подушка, М Т 1995. Использование полуколичественной ПЦР для оценки начала и лечения инфекции Pneumocystis carinii на крысиной модели. Журнал клинической микробиологии , 33 3, 718-24
O’Leary, T.J.; Цай, М.М.; Райт, CF; Подушка, М.Т. 1995. Использование полуколичественной ПЦР для оценки начала и лечения инфекции Pneumocystis carinii на крысиной модели Journal of Clinical Microbiology , 33 3, 718
Andrews, R.P.; Теус, С.А.; Подушка, MT; Уолцер, П.Д. 1994. Сравнение антигенного распознавания основного поверхностного гликопротеина Pneumocystis carinii, выделенного из разных линий крыс Журнал эукариотической микробиологии , 41 5, 72S
Chen, F; Cushion, MT 1994. Использование биолюминесцентного анализа АТФ для оценки жизнеспособности Pneumocystis carinii у крыс. Журнал клинической микробиологии , 32 11, 2791-800
Использование биолюминесцентного анализа АТФ для оценки жизнеспособности Pneumocystis carinii у крыс. Журнал клинической микробиологии , 32 11, 2791-800
Chen,F.; Подушка, М.Т. 1994. Использование биолюминесцентного анализа АТФ для оценки жизнеспособности Pneumocystis carinii у крыс Journal of Clinical Microbiology , 32 11, 2791
Chen, F.; Подушка, М.Т. 1994. Использование флуоресцентных зондов для исследования метаболического состояния митохондрий Pneumocystis carinii Журнал эукариотической микробиологии , 41 5, 79S
Cushion, M T; Хармсен, А; Мацумото, Ю; Стрингер, Дж. Р.; Уэйкфилд, AE; Ямада, М. 1994. Последние достижения в биологии Pneumocystis carinii. Журнал медицинской и ветеринарной микологии: выходящее два раза в месяц издание Международного общества микологии человека и животных , 32 Suppl 1, 217-28
Cushion, M.T.; Хармсен, А.; Мацумото, Ю.; Стрингер, младший; Уэйкфилд, AE; Ямада, М. 1994. Последние достижения в биологии Pneumocystis carinii 9. 0003 Журнал медицинской и ветеринарной микологии: выходящее два раза в месяц издание Международного общества микологии человека и животных , 32 Suppl 1, 217
0003 Журнал медицинской и ветеринарной микологии: выходящее два раза в месяц издание Международного общества микологии человека и животных , 32 Suppl 1, 217
De Stefano, J A; Фой, Дж. М.; Салливан, Д. В.; Доус, С. М.; Подушка, М Т; Бэбкок, Г. Ф.; Слейт, Р. Г.; ван Халбик, Х.; Walzer, PD 1994. Фракционирование стадий развития Pneumocystis carinii с помощью противоточной центробежной элютриации и последовательной фильтрации. Исследования в области паразитологии , 80 1, 1-9
De Stefano, J.A.; Фой, Дж. М.; Салливан, Д. У.; Доус, С.М.; Подушка, MT; Бэбкок, Г.Ф.; Слейт, Р.Г.; ван Халбик, Х.; Уолцер, П.Д. 1994. Фракционирование стадий развития Pneumocystis carinii противоточным центробежным отмыванием и последовательной фильтрацией Исследование паразитологии , 80 1, 1
Langreth, S.G.; Ким, GM; Подушка, М.Т. 1994. Оценка процедур, используемых для выделения Pneumocystis carinii из инфицированных легких крыс The Journal of eukaryotic microbiology , 415, 95S
Settnes, O. P.; Нильсен, П.Б.; Букала, Р.; Линке, М.Дж.; Подушка, М.Т. 1994. Обследование птиц в Дании на наличие Pneumocystis carinii 9.0003 Болезни птиц , 38 1, 1
P.; Нильсен, П.Б.; Букала, Р.; Линке, М.Дж.; Подушка, М.Т. 1994. Обследование птиц в Дании на наличие Pneumocystis carinii 9.0003 Болезни птиц , 38 1, 1
Смулян А.Г.; Линке, М.Дж.; Подушка, М Т; Боуман, Р. П.; Рама, П Т; Дон, М.Н.; Белый, М Л; Walzer, PD 1994. Анализ содержания, жизнеспособности и антигенов Pneumocystis carinii в жидкости бронхоальвеолярного лаважа у больных СПИДом с пневмоцистозом: корреляция с тяжестью заболевания. AIDS (Лондон, Англия) , 8 11, 1555-62
Смулиан, А.Г.; Линке, М.Дж.; Подушка, MT; Боуман, Р.П.; Рама, П.Т.; Дон, М.Н.; Белый, мл.; Уолцер, П.Д. 1994. Анализ содержания, жизнеспособности и антигенов Pneumocystis carinii в жидкости бронхоальвеолярного лаважа у больных СПИДом с пневмоцистозом: корреляция с тяжестью заболевания AIDS (Лондон, Англия) , 8 11, 1555
Vasquez, J.; Смулян, А.Г.; Линке, М.Дж.; Подушка, М.Т. 1994. Антигенные различия среди генетически различных типов Pneumocystis carinii крысиного происхождения Журнал эукариотической микробиологии , 415, 114S
Weinberg, G. A.; Дайкстра, CC; Дюрант, П.Дж.; Подушка, М.Т. 1994. Хромосомная локализация 20 генов в пяти различных кариотипических формах геля пульсирующего поля крыс Pneumocystis carinii The Journal of eukaryotic microbiology , 415, 117S
A.; Дайкстра, CC; Дюрант, П.Дж.; Подушка, М.Т. 1994. Хромосомная локализация 20 генов в пяти различных кариотипических формах геля пульсирующего поля крыс Pneumocystis carinii The Journal of eukaryotic microbiology , 415, 117S
Weinberg, G.A.; О’Гара, М.Дж.; Подушка, М.Т. 1994. Коинфекция крыс генетически разнообразными формами Pneumocystis carinii, продемонстрированная полиморфизмом гена инозинмонофосфатдегидрогеназы P. carinii The Journal of eukaryotic microbiology , 415, 119S
Cushion, MT; Каселис, М; Стрингер, S L; Стрингер, JR 1993. Генетическая стабильность и разнообразие Pneumocystis carinii, заражающих колонии крыс. Инфекция и иммунитет , 61 11, 4801-13
Подушка, М Т; Чжан, Дж.; Каселис, М; Джунтоли, Д.; Стрингер, S L; Стрингер, Дж. Р. 1993. Доказательства двух генетических вариантов Pneumocystis carinii, коинфицирующих лабораторных крыс. Журнал клинической микробиологии , 31 5, 1217-23
Cushion, M.T.; Каселис, М. ; Стрингер, С.Л.; Стрингер, Дж.Р. 1993. Генетическая стабильность и разнообразие Pneumocystis carinii, заражающих колонии крыс Инфекция и иммунитет , 61 11, 4801
; Стрингер, С.Л.; Стрингер, Дж.Р. 1993. Генетическая стабильность и разнообразие Pneumocystis carinii, заражающих колонии крыс Инфекция и иммунитет , 61 11, 4801
Cushion,M.T.; Чжан, Дж.; Каселис, М.; Джунтоли, Д.; Стрингер, С.Л.; Стрингер, Дж.Р. 1993. Доказательства двух генетических вариантов Pneumocystis carinii, коинфицирующих лабораторных крыс Journal of Clinical Microbiology , 31 5, 1217
Stringer, J.R.; Стрингер, С.Л.; Чжан, Дж.; Боуман Р.; Смулян, А.Г.; Подушка, М.Т. 1993. Молекулярно-генетические отличия Pneumocystis carinii от крыс и людей Журнал эукариотической микробиологии , 40 6, 733
Чжан, Дж.; Подушка, М Т; Стрингер, Дж. Р. 1993. Молекулярная характеристика нового повторяющегося элемента из Pneumocystis carinii крыс. Журнал клинической микробиологии , 31 2, 244-8
Zhang,J.; Подушка, MT; Стрингер, Дж.Р. 1993. Молекулярная характеристика нового повторяющегося элемента Pneumocystis carinii от крыс Журнал клинической микробиологии , 312, 244
De Stefano, JA; Тринкль, L S; Уолцер, П. Д.; Подушка, М Т 1992. Проточный цитометрический анализ связывания лектина с поверхностными углеводами Pneumocystis carinii. Журнал паразитологии , 78 2, 271-80
Д.; Подушка, М Т 1992. Проточный цитометрический анализ связывания лектина с поверхностными углеводами Pneumocystis carinii. Журнал паразитологии , 78 2, 271-80
De Stefano, J.A.; Тринкль, Л.С.; Уолцер, PD; Подушка, М.Т. 1992. Проточный цитометрический анализ связывания лектина с поверхностными углеводами Pneumocystis carinii The Journal of parasitology , 782, 271
Stringer, JR; Эдман, Дж. К.; Подушка, М Т; Ричардс, Ф.Ф.; Watanabe, J 1992. Грибковая природа Pneumocystis. Журнал медицинской и ветеринарной микологии: выходящее два раза в месяц издание Международного общества микологии человека и животных , 30 Suppl 1, 271-8
Stringer, J.R.; Эдман, Дж. К.; Подушка, MT; Ричардс, Ф.Ф.; Ватанабэ, Дж. 1992. Грибковая природа Pneumocystis Журнал медицинской и ветеринарной микологии: двухмесячная публикация Международного общества микологии человека и животных , 30 Suppl 1, 271
Cushion, MT; Стрингер, Дж. Р.; Walzer, PD 1991. Клеточная и молекулярная биология Pneumocystis carinii. Международный обзор цитологии , 131, 59-107
Клеточная и молекулярная биология Pneumocystis carinii. Международный обзор цитологии , 131, 59-107
Cushion, M.T.; Линке, М.Дж. 1991. Факторы, влияющие на инфекцию Pneumocystis у крыс с ослабленным иммунитетом The Journal of protozoology , 386, 133S
Cushion, M.T.; Стрингер, младший; Уолцер, П.Д. 1991. Клеточная и молекулярная биология Pneumocystis carinii Международный обзор цитологии , 131, 59
Kaneshiro, E.S.; Ву, Ю.П.; Подушка, М.Т. 1991. Анализы для проверки жизнеспособности Pneumocystis carinii The Journal of protozoology , 386, 85S
Zhang, J.X.; Подушка, MT; Джунтоли, Д.; Хонг, ST; Мид, Дж. К.; Смулян, А.Г.; Стрингер, С.Л.; Стрингер, Дж.Дж. 1991. Анализ генома Pneumocystis carinii Журнал протозоологии , 386, 69S
Cushion, MT; Эббетс, Д. 1990. Рост и метаболизм Pneumocystis carinii в аксенической культуре. Журнал клинической микробиологии , 28 6, 1385-94
Cushion, M T; Хонг, С. Т.; Стил, ЧП; Стрингер, S L; Уолцер, П. Д.; Стрингер, Дж. Р. 1990. Молекулярная биология Pneumocystis carinii. Анналы Нью-Йоркской академии наук , 616, 415-20
Т.; Стил, ЧП; Стрингер, S L; Уолцер, П. Д.; Стрингер, Дж. Р. 1990. Молекулярная биология Pneumocystis carinii. Анналы Нью-Йоркской академии наук , 616, 415-20
Cushion, M.T.; Эббетс, Д. 1990. Рост и метаболизм Pneumocystis carinii в аксенической культуре Журнал клинической микробиологии , 286, 1385
Cushion, M.T.; Хонг, ST; Стил, ЧП; Стрингер, С.Л.; Уолцер, PD; Стрингер, Дж.Р. 1990. Молекулярная биология Pneumocystis carinii Annals of the New York Academy of Sciences , 616, 415
De Stefano, J.A.; Подушка, MT; Пуванесараджа, В.; Уолцер, П.Д. 1990. Анализ стенки кисты Pneumocystis carinii. II. Сахарная композиция Журнал протозоологии , 37 5, 436
Де Стефано, Дж. А.; Подушка, MT; Слейт, Р.Г.; Уолцер, П.Д. 1990. Анализ стенки кисты Pneumocystis carinii. I. Доказательства наличия внешней поверхностной мембраны The Journal of protozoology , 37 5, 428
Hong, S T; Стил, ЧП; Подушка, М Т; Уолцер, П. Д.; Стрингер, S L; Стрингер, Дж. Р. 1990. Кариотипы Pneumocystis carinii. Журнал клинической микробиологии , 28 8, 1785-95
Р. 1990. Кариотипы Pneumocystis carinii. Журнал клинической микробиологии , 28 8, 1785-95
Hong,S.T.; Стил, ЧП; Подушка, MT; Уолцер, PD; Стрингер, С.Л.; Стрингер, Дж.Р. 1990. Кариотипы Pneumocystis carinii Журнал клинической микробиологии , 288, 1785
Cushion, M.T. 1989. Исследования Pneumocystis carinii in vitro The Journal of protozoology , 36 1, 45
Cushion, M.T.; Блейз, Массачусетс; Уолцер, П.Д. 1989. Способ выделения РНК из Pneumocystis carinii The Journal of protozoology , 361, 12S
De Stefano, J.A.; Подушка, MT; Тринкль, Л.С.; Уолцер, П.Д. 1989. Лектины как зонды к поверхностным гликокомплексам Pneumocystis carinii Журнал протозоологии , 36 1, 65S
Канеширо, Э.С.; Подушка, MT; Уолцер, PD; Джаясимхулу, К. 1989. Анализ жирных кислот Pneumocystis The Journal of protozoology , 361, 69S
Linke, MJ; Подушка, М Т; Walzer, PD 1989. Свойства основных антигенов Pneumocystis carinii крысы и человека. Инфекция и иммунитет , 57 5, 1547-55
Инфекция и иммунитет , 57 5, 1547-55
Linke,M.J.; Подушка, MT; Уолцер, П.Д. 1989. Свойства основных антигенов Pneumocystis carinii крысы и человека 9.0003 Инфекция и иммунитет , 57 5, 1547
Ruffolo, J.J.; Подушка, MT; Уолцер, П.Д. 1989. Ультраструктурные наблюдения за стадиями жизненного цикла Pneumocystis carinii The Journal of protozoology , 36 1, 53S
Stringer, S L; Стрингер, Дж. Р.; Блейз, Массачусетс; Уолцер, П. Д.; Cushion, MT 1989. Pneumocystis carinii: последовательность рибосомной РНК предполагает тесную связь с грибами. Экспериментальная паразитология , 68 4, 450-61
Stringer, S.L.; Хадсон, К.; Блейз, Массачусетс; Уолцер, PD; Подушка, MT; Стрингер, Дж.Р. 1989. Последовательность рибосомной РНК Pneumocystis carinii в сравнении с последовательностями четырех грибов предполагает родство с аскомицетами The Journal of protozoology , 36 1, 14S
Stringer, S.L.; Стрингер, младший; Блейз, Массачусетс; Уолцер, PD; Подушка, М. Т. 1989. Pneumocystis carinii: последовательность рибосомной РНК предполагает тесную связь с грибами Experimental parasitology , 684, 450
Т. 1989. Pneumocystis carinii: последовательность рибосомной РНК предполагает тесную связь с грибами Experimental parasitology , 684, 450
Walzer, PD; Cushion, MT 1989. Иммунобиология Pneumocystis carinii. Исследования в области патологии и иммунопатологии , 8 3-4, 127-40
Walzer, P.D.; Подушка, М.Т. 1989. Иммунобиология Pneumocystis carinii Патологические и иммунопатологические исследования , 8 3-4, 127
Cushion, MT; ДеСтефано, Дж. А.; Walzer, PD 1988. Pneumocystis carinii: поверхностно-реактивные углеводы, обнаруженные с помощью лектиновых зондов. Экспериментальная паразитология , 67 2, 137-47
Подушка, М Т; Руффоло, Дж. Дж.; Walzer, PD 1988. Анализ стадий развития Pneumocystis carinii in vitro. Лабораторные исследования; журнал технических методов и патологии , 58 3, 324-31
Подушка, M.T.; ДеСтефано, Дж. А.; Уолцер, П.Д. 1988. Pneumocystis carinii: поверхностно-реактивные углеводы, обнаруженные с помощью лектиновых зондов Experimental Parasitology , 67 2, 137
Cushion, M. T.; Руффоло, Дж. Дж.; Уолцер, П.Д. 1988. Анализ стадий развития Pneumocystis carinii, in vitro Лабораторное исследование; журнал технических методов и патологии , 58 3, 324
T.; Руффоло, Дж. Дж.; Уолцер, П.Д. 1988. Анализ стадий развития Pneumocystis carinii, in vitro Лабораторное исследование; журнал технических методов и патологии , 58 3, 324
Walzer, PD; Ким, СК; Фой, Дж. М.; Линке, М.Дж.; Подушка, М Т 1988. Ингибиторы синтеза фолиевой кислоты в лечении экспериментальной пневмоцистной пневмонии, вызванной Pneumocystis carinii. Противомикробные средства и химиотерапия , 32 1, 96-103
Walzer, PD; Ким, СК; Фой, Дж.; Линке, М.Дж.; Cushion, MT 1988. Катионные антитрипаносомные и другие антимикробные агенты в терапии экспериментальной пневмонии, вызванной Pneumocystis carinii. Противомикробные средства и химиотерапия , 32 6, 896-905
Walzer, P.D.; Ким, СК; Фой, Дж.; Линке, М.Дж.; Подушка, М.Т. 1988. Катионные антитрипаносомные и другие противомикробные средства в терапии экспериментальной пневмоцистной пневмонии Pneumocystis carinii Противомикробные средства и химиотерапия , 32 6, 896
Walzer, P. D.; Ким, СК; Фой, Дж. М.; Линке, М.Дж.; Подушка, М.Т. 1988. Ингибиторы синтеза фолиевой кислоты при лечении экспериментальной пневмоцистной пневмонии Pneumocystis carinii Антимикробные агенты и химиотерапия , 32 1, 96
D.; Ким, СК; Фой, Дж. М.; Линке, М.Дж.; Подушка, М.Т. 1988. Ингибиторы синтеза фолиевой кислоты при лечении экспериментальной пневмоцистной пневмонии Pneumocystis carinii Антимикробные агенты и химиотерапия , 32 1, 96
Kim, CK; Фой, Дж. М.; Подушка, М Т; Стэнфорт, Д.; Линке, М.Дж.; Хендрикс, Х.Л.; Уолцер, PD 1987. Сравнение гистологических и количественных методов оценки терапии экспериментальной пневмоцистной пневмонии, вызванной Pneumocystis carinii. Противомикробные средства и химиотерапия , 31 2, 197-201
Kim, C.K.; Фой, Дж. М.; Подушка, MT; Стэнфорт, Д.; Линке, М.Дж.; Хендрикс, HL; Уолцер, П.Д. 1987. Сравнение гистологических и количественных методов в оценке терапии экспериментальной пневмоцистной пневмонии Pneumocystis carinii Антимикробные агенты и химиотерапия , 31 2, 197
Уолцер, П. Д.; Подушка, М Т; Юранек, Д.; Стены, К; Армстронг, Д.; Золото, Дж; Янг, Л.С.; Песанти, Э; Грейвс, округ Колумбия; Ivey, MB 1987. Серология и P. carinii. Сундук , 91 6, 935-6
carinii. Сундук , 91 6, 935-6
Walzer, PD; Стэнфорт, Д.; Линке, М.Дж.; Cushion, MT 1987. Pneumocystis carinii: иммуноблоттинг и иммунофлуоресцентный анализ сывороточных антител во время экспериментальной крысиной инфекции и выздоровления. Экспериментальная паразитология , 63 3, 319-28
Walzer, P.D.; Подушка, MT; Джуранек, Д.; Уоллс, К.; Армстронг, Д.; Голд, Дж.; Янг, Л.С.; Песанти, Э.; Грейвс, округ Колумбия; Айви, М.Б. 1987. Серология и P. carinii Chest , 91 6, 935
Walzer, P.D.; Стэнфорт, Д.; Линке, М.Дж.; Подушка, М.Т. 1987. Pneumocystis carinii: иммуноблоттинг и иммунофлуоресцентный анализ сывороточных антител во время экспериментального заражения крыс и восстановления Экспериментальная паразитология , 633, 319
Ruffolo, JJ; Подушка, М Т; Walzer, PD 1986. Методы исследования Pneumocystis carinii в свежих образцах. Журнал клинической микробиологии , 23 1, 17-21
Руффоло, Джей Джей; Подушка, MT; Уолцер, П. Д. 1986. Методы исследования Pneumocystis carinii в свежих образцах Журнал клинической микробиологии , 23 1, 17
Д. 1986. Методы исследования Pneumocystis carinii в свежих образцах Журнал клинической микробиологии , 23 1, 17
Cushion, MT; Руффоло, Дж. Дж.; Линке, М.Дж.; Walzer, PD 1985. Pneumocystis carinii: параметры роста и оценки в линиях клеток человека A549 и WI-38 VA13. Экспериментальная паразитология , 60 1, 43-54
Подушка, М Т; Стэнфорт, Д.; Линке, М.Дж.; Walzer, PD 1985. Метод тестирования чувствительности Pneumocystis carinii к противомикробным агентам in vitro. Противомикробные препараты и химиотерапия , 28 6, 796-801
Cushion, M.T.; Руффоло, Дж. Дж.; Линке, М.Дж.; Уолцер, П.Д. 1985. Pneumocystis carinii: переменные роста и оценки в линиях клеток человека A549 и WI-38 VA13 Экспериментальная паразитология , 601, 43
Cushion, M.T.; Стэнфорт, Д.; Линке, М.Дж.; Уолцер, П.Д. 1985. Метод тестирования чувствительности Pneumocystis carinii к антимикробным агентам in vitro Антимикробные агенты и химиотерапия , 28 6, 796
Подушка, M T; Walzer, PD 1984. Рост и серийное пассирование Pneumocystis carinii в клеточной линии A549. Инфекция и иммунитет , 44 2, 245-51
Рост и серийное пассирование Pneumocystis carinii в клеточной линии A549. Инфекция и иммунитет , 44 2, 245-51
Подушка, М Т; Walzer, PD 1984. Культивирование Pneumocystis carinii в клеточных линиях легких. Журнал инфекционных болезней , 149 4, 644
Cushion, M.T.; Уолцер, П.Д. 1984. Рост и серийное пассирование Pneumocystis carinii в клеточной линии А549 Инфекция и иммунитет , 44 2, 245
Подушка, M.T.; Уолцер, П.Д. 1984. Культивирование Pneumocystis carinii в клеточных линиях легких Журнал инфекционных заболеваний , 1494, 644
Walzer, PD; Лабин, М.; Редингтон, Т.Дж.; Cushion, MT 1984. Предрасполагающие факторы пневмоцистной пневмонии: воздействие тетрациклина, белковой недостаточности и кортикостероидов на хозяев. Инфекция и иммунитет , 46 3, 747-53
Walzer, PD; Лабин, М.; Редингтон, Т.Дж.; Cushion, MT 1984. Изменения лимфоцитов при хроническом введении и отмене кортикостероидов: связь с пневмонией, вызванной Pneumocystis carinii. Journal of Immunology (Балтимор, Мэриленд: 1950) , 133 5, 2502-8
Journal of Immunology (Балтимор, Мэриленд: 1950) , 133 5, 2502-8
Walzer, P.D.; Лабин, М.; Редингтон, Т.Дж.; Подушка, М.Т. 1984. Предрасполагающие факторы пневмоцистной пневмонии: влияние тетрациклина, белковой недостаточности и кортикостероидов на хозяев Инфекция и иммунитет , 46 3, 747
Walzer, P.D.; Лабин, М.; Редингтон, Т.Дж.; Подушка, М.Т. 1984. Изменения лимфоцитов при хроническом приеме и отмене кортикостероидов: связь с пневмонией Pneumocystis carinii Journal of Immunology (Балтимор, штат Мэриленд: 1950) , 133 5, 2502
Gomes A.; Ферраз Р.; Фикер Л.; Коллинз М.С.; Пруденсио К.; Подушка М.Т.; Тейшейра С.; Гомеш П. 01-11-2018. Аналоги хлорохина как ведущие против легочных патогенов Pneumocystis Противомикробные агенты и химиотерапия , 62 11,
Luraschi A.; Ричард С.; Алмейда Дж. М.; Паньи М.; Подушка М.Т.; Хаузер П.М. 07-01-2019. Анализы экспрессии и иммунного окрашивания позволяют предположить, что первичный гомоталлиз пневмоцистиса включает троф 9. 0003 mBio , 10 4,
0003 mBio , 10 4,
Мацеевская Д.; Забиски Дж.; Резлер М.; Камерчак П.; Коллинз М.; Фикер Л.; Подушка М. 01-01-2017. Разработка высокоактивных анти-: Pneumocystis bisbenzamidines: Insight in the effect of sele MedChemComm , 8 10, 2003-2011
Andrews, RP; Теус, SA; Подушка, М Т; Уолцер, П.Д. Сравнение антигенного распознавания основного поверхностного гликопротеина Pneumocystis carinii, выделенного из разных линий крыс. Журнал эукариотической микробиологии , 41 5, 72S
Арнольд, Дж.; Подушка, М Т . Построение физической карты генома Pneumocystis. Журнал эукариотической микробиологии , 44 6, 8S
Chen, F; Подушка, М Т . Использование флуоресцентных зондов для исследования метаболического состояния митохондрий Pneumocystis carinii. Журнал эукариотической микробиологии , 41 5, 79S
Cushion, M T . Резюме исследования Pneumocystis, представленное на 6-м Международном семинаре по оппортунистическим протистам. Журнал эукариотической микробиологии , 46 5, 85S-88S
Cushion, M T . Исследования Pneumocystis carinii in vitro. Журнал протозоологии , 36 1, 45-52
Исследования Pneumocystis carinii in vitro. Журнал протозоологии , 36 1, 45-52
Подушка, М Т; Арнольд, Дж. Предложение по проекту генома Pneumocystis. Журнал эукариотической микробиологии , 44 6, 7S
Cushion, MT; Блейз, Массачусетс; Уолцер, П.Д. Способ выделения РНК из Pneumocystis carinii. Журнал протозоологии , 36 1, 12С-14С
Подушка, M T; Линке, М.Дж. Факторы, влияющие на инфекцию Pneumocystis у крыс с ослабленным иммунитетом. Журнал протозоологии , 38 6, 133S-135S
Подушка, М Т; Линке, М.Дж.; Коллинз, М; Кили, С.П.; Стрингер, Дж. Р. Минимальное количество Pneumocystis carinii f. сп. carinii, необходимое для установления инфекций, очень мало. Журнал эукариотической микробиологии , 46 5, 111S
Cushion, MT; Орр, С. Кинетика двух генетически различных популяций Pneumocystis carinii в колониях крыс. Журнал эукариотической микробиологии , 43 5, 46S
Cushion, M T; Орр, С; Арнольд, Дж. Взаимодействия между двумя популяциями Pneumocystis в пределах одного хозяина. Журнал эукариотической микробиологии , 44 6, 9S
Взаимодействия между двумя популяциями Pneumocystis в пределах одного хозяина. Журнал эукариотической микробиологии , 44 6, 9S
Cushion, MT; Уолцер, П.Д. Заключительные комментарии: ретроспективная перспектива 4-го Международного семинара по Pneumocystis. Журнал эукариотической микробиологии , 43 5, 60S-61S
Cushion, Melanie T . Сравнительная геномика Pneumocystis carinii с другими простейшими: влияние на образ жизни. Журнал эукариотической микробиологии , 51 1, 30-7
Cushion, Melanie T; Кили, Скотт П.; Стрингер, Джеймс Р. Молекулярно-фенотипическое описание Pneumocystis wakefieldiae sp. nov., новый вид у крыс. Mycologia , 96 3, 429-38
De Stefano, J A; Подушка, М Т; Пуванесараджа, В.; Уолцер, П.Д. Анализ стенки кисты Pneumocystis carinii. II. Сахарный состав. Журнал протозоологии , 37 5, 436-41
Де Стефано, Дж. А.; Подушка, М Т; Слейт, Р. Г.; Уолцер, П.Д. Анализ стенки кисты Pneumocystis carinii. I. Доказательства наличия наружной поверхностной мембраны. Журнал протозоологии , 37 5, 428-35
I. Доказательства наличия наружной поверхностной мембраны. Журнал протозоологии , 37 5, 428-35
Де Стефано, Дж. А.; Подушка, М Т; Тринкль, L S; Уолцер, П.Д. Лектины как зонды к поверхностным гликокомплексам Pneumocystis carinii. Журнал протозоологии , 36 1, 65S-66S
Icenhour, CR; Арнольд, Дж.; Подушка, М Т . Взаимодействие двух популяций Pneumocystis carinii в легких крыс. Журнал эукариотической микробиологии , 46 5, 107S-108S
Канеширо, Э.С.; Коллинз, М; Подушка, М Т . Влияние ингибиторов стеролов на содержание АТФ в Pneumocystis carinii. Журнал эукариотической микробиологии , 46 5, 142S-143S
Kaneshiro, E S; Подушка, М Т; Уолцер, П. Д.; Джаясимхулу, К. Анализ жирных кислот Pneumocystis. Журнал протозоологии , 36 1, 69S-72S
Канеширо, Э.С.; Ву, Ю П; Подушка, М Т . Анализы для проверки жизнеспособности Pneumocystis carinii. Журнал протозоологии , 38 6, 85S-87S
Канеширо, Эдна С; Подушка, Мелани Т. ; Марчиано-Кабрал, Франсин; Вайс, Луи М.; Сяо, Лихуа. Основные моменты и итоги 11-го Международного семинара по оппортунистическим протистам. Журнал эукариотической микробиологии , 58 1, 1-6
; Марчиано-Кабрал, Франсин; Вайс, Луи М.; Сяо, Лихуа. Основные моменты и итоги 11-го Международного семинара по оппортунистическим протистам. Журнал эукариотической микробиологии , 58 1, 1-6
Keely, S P; Подушка, М Т; Стрингер, Дж. Р. Определение максимальной частоты генетических перестроек, связанных с вариацией поверхностного антигена Pneumocystis carinii. Журнал эукариотической микробиологии , 46 5, 128S
Keely, S P; Подушка, М Т; Стрингер, Дж. Р. Стабильность четырех генетических локусов у Pneumocystis carinii sp. ф. карины. Журнал эукариотической микробиологии , 43 5, 49S
Лангрет, С. Г.; Ким, ГМ; Подушка, М Т . Оценка процедур, используемых для выделения Pneumocystis carinii из инфицированных легких крыс. Журнал эукариотической микробиологии , 41 5, 95S-96S
Redhead, Scott A; Подушка, Мелани Т.; Френкель, Джейкоб К.; Стрингер, Джеймс Р. Pneumocystis и Trypanosoma cruzi: номенклатура и типизация. Журнал эукариотической микробиологии , 53 1, 2-11
Ruffolo, J J; Подушка, М Т; Уолцер, П. Д. Ультраструктурные наблюдения за стадиями жизненного цикла Pneumocystis carinii. Журнал протозоологии , 36 1, 53S-54S
Д. Ультраструктурные наблюдения за стадиями жизненного цикла Pneumocystis carinii. Журнал протозоологии , 36 1, 53S-54S
Settnes, O P; Нильсен, П.Б.; Букала, Р.; Линке, М.Дж.; Подушка, М Т . Обследование птиц в Дании на наличие Pneumocystis carinii. Болезни птиц , 38 1, 1-10
Sloand, E; Логон, Б; Армстронг, М.; Бартлетт, М.С.; Блюменфельд, Вт; Подушка, М; Калица, А; Ковач, Дж. А.; Мартин, В.; Питт, Э. Вызов культуры Pneumocystis carinii. Журнал эукариотической микробиологии , 40 2, 188-95
Стрингер, JR; Стрингер, S L; Чжан, Дж.; Боуман, Р.; Смулян, А. Г.; Подушка, М Т . Молекулярно-генетические отличия Pneumocystis carinii от крыс и человека. Журнал эукариотической микробиологии , 406, 733-41
Stringer, JR; Уэйкфилд, AE; Подушка, М Т; Дей-Кас, Э. Таксономия и номенклатура Pneumocystis: обновление. Журнал эукариотической микробиологии , 44 6, 5S-6S
Stringer, S L; Хадсон, К.; Блейз, Массачусетс; Уолцер, П. Д.; Подушка, М Т; Стрингер, Дж. Р. Последовательность рибосомной РНК Pneumocystis carinii по сравнению с последовательностями четырех грибов предполагает родство с аскомицетами. Журнал протозоологии , 36 1, 14S-16S
Д.; Подушка, М Т; Стрингер, Дж. Р. Последовательность рибосомной РНК Pneumocystis carinii по сравнению с последовательностями четырех грибов предполагает родство с аскомицетами. Журнал протозоологии , 36 1, 14S-16S
Vasquez, J; Смулян, А. Г.; Линке, М.Дж.; Подушка, М Т . Антигенные различия между генетически различными типами Pneumocystis carinii крысиного происхождения. Журнал эукариотической микробиологии , 41 5, 114S
Weinberg, GA; Дыкстра, С С; Дюрант, П.Дж.; Подушка, М Т . Хромосомная локализация 20 генов пяти различных кариотипических форм геля пульсирующего поля крыс Pneumocystis carinii. Журнал эукариотической микробиологии , 41 5, 117S
Вайнберг, Г. А.; О’Гара, MJ; Подушка, М Т . Коинфицирование крыс генетически разнообразными формами Pneumocystis carinii, демонстрируемое полиморфизмом гена инозинмонофосфатдегидрогеназы P. carinii. Журнал эукариотической микробиологии , 41 5, 119S
Zhang, J X; Подушка, М Т; Джунтоли, Д. ; Хонг, С. Т.; Мид, Дж. К.; Смулян, А. Г.; Стрингер, S L; Стрингер, Дж.Дж. Анализ генома Pneumocystis carinii. Журнал протозоологии , 38 6, 69S
; Хонг, С. Т.; Мид, Дж. К.; Смулян, А. Г.; Стрингер, S L; Стрингер, Дж.Дж. Анализ генома Pneumocystis carinii. Журнал протозоологии , 38 6, 69S
SOX9 модулирует экспрессию ключевых факторов транскрипции, необходимых для развития сердечного клапана | Разработка
Пропустить пункт назначения
ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ| 15 декабря 2015 г.
Виктория С. Гарсайд,
Ребекка Каллум,
Оливия Алдер,
Дафна Ю. Лу,
Райан Вандер Верфф,
Михаил Беленький,
Юнджун Чжао,
Стивен Дж. М. Джонс,
М. Джонс,
Марко А. Марра,
Т. Майкл Андерхилл,
Памела А. Без капюшона
Информация об авторе и статье
*Автор для переписки ([email protected])
Конкурирующие интересы
Авторы заявляют об отсутствии конкурирующих или финансовых интересов.
Полученный: 13 апр 2015
Принято: 28 октября 2015 г.
Номер в сети: 1477-9129
Номер для печати: 0950-1991
© 2015. Опубликовано The Company of Biologist Ltd
2015
Развитие (2015) 142 (24): 4340–4350.
https://doi.org/10.1242/dev.125252
История статьи
Получено:
13 апреля 2015 г.
Принято:
28 октября 2015 г.
- Разделенный экран
- Просмотры
- Содержание статьи
- Рисунки и таблицы
- Видео
- Аудио
- Дополнительные данные
- Экспертная оценка
- PDF ПДФ+СИ
- Значок версии статьи Версии
- Версия записи
15 декабря 2015 г.
- Принятая рукопись 01 января 2015 г.
- Версия записи
15 декабря 2015 г.
- Делиться
- MailTo
- Твиттер
- Инструменты
Получить разрешения
Иконка Цитировать Цитировать
- Поиск по сайту

Citation
Виктория С. Гарсайд, Ребекка Каллум, Оливия Алдер, Дафна Ю. Лу, Райан Вандер Верфф, Михаил Биленки, Юнджун Чжао, Стивен Дж. М. Джонс, Марко А. Марра, Т. Майкл Андерхилл, Памела А. Худлесс; SOX9модулирует экспрессию ключевых факторов транскрипции, необходимых для развития сердечного клапана. Развитие 15 декабря 2015 г.; 142 (24): 4340–4350. doi: https://doi.org/10.1242/dev.125252
Гарсайд, Ребекка Каллум, Оливия Алдер, Дафна Ю. Лу, Райан Вандер Верфф, Михаил Биленки, Юнджун Чжао, Стивен Дж. М. Джонс, Марко А. Марра, Т. Майкл Андерхилл, Памела А. Худлесс; SOX9модулирует экспрессию ключевых факторов транскрипции, необходимых для развития сердечного клапана. Развитие 15 декабря 2015 г.; 142 (24): 4340–4350. doi: https://doi.org/10.1242/dev.125252
Скачать файл цитаты:
- Рис (Зотеро)
- Диспетчер ссылок
- EasyBib
- Подставки для книг
- Менделей
- Бумаги
- Конечная примечание
- RefWorks
- Бибтекс
Расширенный поиск
Формирование сердечного клапана начинается, когда эндотелиальные клетки сердца трансформируются в мезенхиму и заселяют сердечные подушки. Фактор транскрипции SOX9 в высокой степени экспрессируется в мезенхиме кардиальной подушки и необходим для развития сердечных клапанов. Потеря Sox9 в мезенхиме кардиальной подушки мыши изменяет пролиферацию клеток, выживание эмбриона и формирование клапана. Несмотря на эту важную роль, мало что известно о том, как SOX9 регулирует формирование сердечного клапана или его транскрипционные мишени. Поэтому мы нанесли на карту предполагаемые сайты связывания SOX9 с помощью ChIP-Seq в сердечных клапанах E12.5, стадии, на которой мезенхима клапана активно пролиферирует и инициирует дифференцировку. Было показано, что эмбриональные сердечные клапаны экспрессируют большое количество генов, связанных с хондрогенезом, включая несколько белков внеклеточного матрикса и факторы транскрипции, которые регулируют хондрогенез. Поэтому мы сравнили области предполагаемого SOX9.Связывание ДНК между сердечными клапанами E12.5 и зачатками конечностей E12.5. Мы идентифицировали контекстно-зависимые и контекстно-независимые SOX9-взаимодействующие области по всему геному. Анализ независимого от контекста связывания SOX9 предполагает обширную роль SOX9 в тканях в регуляции связанных с пролиферацией генов, включая ключевые компоненты комплекса AP-1.
Потеря Sox9 в мезенхиме кардиальной подушки мыши изменяет пролиферацию клеток, выживание эмбриона и формирование клапана. Несмотря на эту важную роль, мало что известно о том, как SOX9 регулирует формирование сердечного клапана или его транскрипционные мишени. Поэтому мы нанесли на карту предполагаемые сайты связывания SOX9 с помощью ChIP-Seq в сердечных клапанах E12.5, стадии, на которой мезенхима клапана активно пролиферирует и инициирует дифференцировку. Было показано, что эмбриональные сердечные клапаны экспрессируют большое количество генов, связанных с хондрогенезом, включая несколько белков внеклеточного матрикса и факторы транскрипции, которые регулируют хондрогенез. Поэтому мы сравнили области предполагаемого SOX9.Связывание ДНК между сердечными клапанами E12.5 и зачатками конечностей E12.5. Мы идентифицировали контекстно-зависимые и контекстно-независимые SOX9-взаимодействующие области по всему геному. Анализ независимого от контекста связывания SOX9 предполагает обширную роль SOX9 в тканях в регуляции связанных с пролиферацией генов, включая ключевые компоненты комплекса AP-1. Интегративный анализ тканеспецифических областей, взаимодействующих с SOX9, и профилей экспрессии генов на клапанах сердца с дефицитом Sox9 продемонстрировал, что SOX9контролирует экспрессию нескольких факторов транскрипции с ранее идентифицированными ролями в развитии сердечных клапанов, включая Twist1 , Sox4 , Mecom и Pitx2 . Вместе наши данные идентифицируют SOX9-скоординированные иерархии транскрипции, которые контролируют пролиферацию и дифференцировку клеток во время формирования клапана.
Интегративный анализ тканеспецифических областей, взаимодействующих с SOX9, и профилей экспрессии генов на клапанах сердца с дефицитом Sox9 продемонстрировал, что SOX9контролирует экспрессию нескольких факторов транскрипции с ранее идентифицированными ролями в развитии сердечных клапанов, включая Twist1 , Sox4 , Mecom и Pitx2 . Вместе наши данные идентифицируют SOX9-скоординированные иерархии транскрипции, которые контролируют пролиферацию и дифференцировку клеток во время формирования клапана.
Ключевые слова:
Транскрипционные сети, SOX9, Сердечные клапаны, Распространение, Транскрипционный фактор, конечность, Чип-Seq, геном, мышь, Эмбриогенез
Треть сердечно-сосудистых врожденных пороков связана с аномальным формированием сердечных клапанов, и клапанная недостаточность может привести к повышенной восприимчивости к заболеваниям в более позднем возрасте. Клапаны сердца развиваются в двух сужениях в эмбриональной сердечной трубке: между предсердиями и желудочками, известными как атриовентрикулярный канал (AVC), и в выводном тракте (OFT), который образует основание аорты и легочного ствола (обзор de Вламинг и др., 2012). Эти области формируют митральный и трикуспидальный клапаны, аортальный и легочный клапаны соответственно (rev. Person et al., 2005). Начиная с эмбрионального дня (E) 9.5 у мышей эпителиально-мезенхимальный переход (ЭМП) происходит в эндокардиальных клетках АВК, частично в ответ на активацию передачи сигналов Notch (Timmerman et al., 2004), и приводит к их трансформации в мигрирующие клетки. Эти клетки проникают во внеклеточный матрикс (ECM), который разделяет эндокард и миокард, и заселяют его, образуя сердечные подушки, предшественники клапанов и перегородок (rev. Person et al., 2005). Эти мезенхимальные клетки будут пролиферировать, дифференцироваться и реконструироваться, чтобы в конечном итоге сформировать зрелые, тонкие, нежные створки клапана (rev.
Клапаны сердца развиваются в двух сужениях в эмбриональной сердечной трубке: между предсердиями и желудочками, известными как атриовентрикулярный канал (AVC), и в выводном тракте (OFT), который образует основание аорты и легочного ствола (обзор de Вламинг и др., 2012). Эти области формируют митральный и трикуспидальный клапаны, аортальный и легочный клапаны соответственно (rev. Person et al., 2005). Начиная с эмбрионального дня (E) 9.5 у мышей эпителиально-мезенхимальный переход (ЭМП) происходит в эндокардиальных клетках АВК, частично в ответ на активацию передачи сигналов Notch (Timmerman et al., 2004), и приводит к их трансформации в мигрирующие клетки. Эти клетки проникают во внеклеточный матрикс (ECM), который разделяет эндокард и миокард, и заселяют его, образуя сердечные подушки, предшественники клапанов и перегородок (rev. Person et al., 2005). Эти мезенхимальные клетки будут пролиферировать, дифференцироваться и реконструироваться, чтобы в конечном итоге сформировать зрелые, тонкие, нежные створки клапана (rev. de Vlaming et al., 2012).
de Vlaming et al., 2012).
Факторы транскрипции (TF) необходимы для точной координации спецификации и дифференцировки клонов и, в конечном счете, для контроля клеточной идентичности. В сердечном клапане несколько TF вносят свой вклад в эту роль, но как эти TF координируются, чтобы регулировать формирование клапана, неизвестно. SOX (блок SRY-типа) 9, высокоподвижная группа (HMG)-box TF, в высокой степени экспрессируется во вновь образованной мезенхиме развивающихся сердечных клапанов (Montero et al., 2002), и его экспрессия находится непосредственно после передачи сигналов Notch. (Чанг и др., 2014). У мышей SOX9играет существенную роль в развитии сердечного клапана (Lincoln et al., 2007): потеря SOX9 на ранней стадии формирования клапана приводит к гипопластическим эндокардиальным подушкам, снижению пролиферации и изменению отложения ВКМ, тогда как на более поздних стадиях развития клапана потеря SOX9 вызывает аномальные Паттернирование внеклеточного матрикса, потеря связанных с хрящом белков и утолщение клапанов (Lincoln et al. , 2007). Отсутствие SOX9 в любой момент развития сердечного клапана приводит к летальному исходу эмбриона. У человека гаплонедостаточные мутации в гене SOX9locus вызывают кампомелическую дисплазию (Wagner et al., 1994), при которой наблюдаются скелетные аномалии, такие как искривление длинных костей, смена пола и множественные дефекты органов, включая пороки поджелудочной железы и сердца, которые часто приводят к неонатальной смерти (обзор Джейн и Сен, 2014).
, 2007). Отсутствие SOX9 в любой момент развития сердечного клапана приводит к летальному исходу эмбриона. У человека гаплонедостаточные мутации в гене SOX9locus вызывают кампомелическую дисплазию (Wagner et al., 1994), при которой наблюдаются скелетные аномалии, такие как искривление длинных костей, смена пола и множественные дефекты органов, включая пороки поджелудочной железы и сердца, которые часто приводят к неонатальной смерти (обзор Джейн и Сен, 2014).
Эмбриональные сердечные клапаны экспрессируют большое количество генов, связанных с хондрогенезом, включая гены белков внеклеточного матрикса и ряд общих ключевых TF, таких как SOX9, TWIST1 и NFATC1 (Chakraborty et al., 2010a). Подобно сердечным клапанам, хрящевые ткани, такие как конечность, дифференцируются от SOX9.-позитивная мезенхима, предполагая, что их транскрипционные программы аналогичны. Во время формирования хряща в развивающихся зачатках конечностей потеря SOX9 перед мезенхимальной конденсацией приводит к агенезии хряща и последующей потере формирования кости (Akiyama et al. , 2002). Зачатки конечностей с делецией SOX9 после начального образования хряща имеют мезенхимальные уплотнения, которые сильно гипопластичны и имеют дефекты дифференцировки и пролиферации хондроцитов (Akiyama et al., 2002). Потеря SOX9в других системах органов также образуются гипопластические органы и дефекты пролиферации (Lincoln et al., 2007; Trowe et al., 2010; Rockich et al., 2013).
, 2002). Зачатки конечностей с делецией SOX9 после начального образования хряща имеют мезенхимальные уплотнения, которые сильно гипопластичны и имеют дефекты дифференцировки и пролиферации хондроцитов (Akiyama et al., 2002). Потеря SOX9в других системах органов также образуются гипопластические органы и дефекты пролиферации (Lincoln et al., 2007; Trowe et al., 2010; Rockich et al., 2013).
Несмотря на существенную роль SOX9, механизмы его действия и его транскрипционные мишени изучены недостаточно. В стволовых клетках волосяных фолликулов взрослых (HF-SCs) гены, связанные с SOX9, необходимы клеткам для поддержания стволовости посредством секретируемых факторов в нише (Kadaja et al., 2014). Недавно глобальные мишени транскрипции SOX9в хондроцитах начали исследовать (Liu and Lefebvre, 2015; Ohba et al., 2015). Однако в конечностях эмбриона было идентифицировано лишь ограниченное число добросовестных in vivo мишеней транскрипции SOX9 (Bell et al., 1997; Bridgewater et al. , 1998), и нет прямых in vivo мишеней транскрипции SOX9. известен в эмбриональных клапанах сердца.
, 1998), и нет прямых in vivo мишеней транскрипции SOX9. известен в эмбриональных клапанах сердца.
Чтобы изучить in vivo функциональную значимость SOX9 в развитии клапанов и конечностей, мы использовали иммунопреципитацию хроматина в сочетании с секвенированием нового поколения (ChIP-Seq) для создания полногеномных профилей предполагаемого SOX9.Сайты связывания ДНК в АВК и конечностях эмбриона мыши E12.5. Мы идентифицировали 2607 и 9092 области пика SOX9, соответствующие 2453 и 5750 потенциальным генам-мишеням в E12.5 AVC и конечности соответственно. Пики SOX9, характерные для многих тканей, подтверждают роль SOX9 в регуляции клеточной пролиферации и специфически идентифицируют белки AP-1 и другие регуляторы клеточного цикла как мишени для взаимодействия SOX9. Более того, в сердечных клапанах мы идентифицировали группу незаменимых ТФ, модулируемых SOX9. Сравнивая эти потенциальные гены-мишени SOX9к транскрипционным изменениям, идентифицированным в данных RNA-Seq из Sox9 -дефицитных AVC, мы продемонстрировали, что SOX9 является необходимым вышестоящим регулятором, необходимым для модулирования уровней экспрессии факторов, необходимых для формирования сердечного клапана.
 , 1997; Bridgewater et al., 1998; Sekiya et al., 2000 г.) (рис. S1C). Чтобы определить сходство в программах транскрипции, инициируемых SOX9, мы сравнили библиотеки SOX9 AVC и ChIP-Seq конечностей и обнаружили 782 пика SOX9, которые были общими (30,0% пиков E12.5 AVC) (рис. 1A), подтверждая, что SOX9 имеет общие регуляторные роли в развитии клапана и конечности. Дополнительные 1825 и 8310 SOX9пики были уникальными в E12.5 AVC и конечности, соответственно, что указывает на то, что SOX9 также имеет тканеспецифические сайты связывания.
, 1997; Bridgewater et al., 1998; Sekiya et al., 2000 г.) (рис. S1C). Чтобы определить сходство в программах транскрипции, инициируемых SOX9, мы сравнили библиотеки SOX9 AVC и ChIP-Seq конечностей и обнаружили 782 пика SOX9, которые были общими (30,0% пиков E12.5 AVC) (рис. 1A), подтверждая, что SOX9 имеет общие регуляторные роли в развитии клапана и конечности. Дополнительные 1825 и 8310 SOX9пики были уникальными в E12.5 AVC и конечности, соответственно, что указывает на то, что SOX9 также имеет тканеспецифические сайты связывания.Рис. 1.
Просмотреть в большом размереЗагрузить слайд
Пики SOX9 часто локализуются в TSS/промоторе в АВК и конечности. (A) Диаграмма Венна геномного расположения пиков показывает, сколько пиков непосредственно перекрываются между библиотеками. (B) Диаграмма Венна, показывающая гены, которые либо являются уникальными мишенями, либо обычно являются мишенями в E12.5 AVC и ChIP-Seq конечностей. Ген с ассоциированным SOX9пик в обеих тканях может иметь пик в одном и том же месте в обеих тканях или в уникальных тканеспецифических областях, все еще нацеленных на один и тот же ген. (C) Модели позиционного веса для мотивов связывания мономера и димера SOX, идентифицированные по данным ChIP-Seq. Высота букв указывает на важность основания для связывания SOX9. (D) Распределение мест пика SOX9 по геному в E12.5 AVC, конечности E12.5 и общих сайтах связывания.
(C) Модели позиционного веса для мотивов связывания мономера и димера SOX, идентифицированные по данным ChIP-Seq. Высота букв указывает на важность основания для связывания SOX9. (D) Распределение мест пика SOX9 по геному в E12.5 AVC, конечности E12.5 и общих сайтах связывания.
Рис. 1.
Посмотреть в большом размереСкачать слайд
SOX9пики часто расположены в TSS/промоторе в AVC и конечности. (A) Диаграмма Венна геномного расположения пиков показывает, сколько пиков непосредственно перекрываются между библиотеками. (B) Диаграмма Венна, показывающая гены, которые либо являются уникальными мишенями, либо обычно являются мишенями в E12.5 AVC и ChIP-Seq конечностей. Ген со связанным пиком SOX9 в обеих тканях может иметь пик в одном и том же месте в обеих тканях или в уникальных тканеспецифических областях, все еще нацеленных на один и тот же ген. (C) Модели позиционного веса для мотивов связывания мономера и димера SOX, идентифицированные по данным ChIP-Seq. Высота буквы указывает на важность основания для SOX9.привязка. (D) Распределение мест пика SOX9 по геному в E12.5 AVC, конечности E12.5 и общих сайтах связывания.
Высота буквы указывает на важность основания для SOX9.привязка. (D) Распределение мест пика SOX9 по геному в E12.5 AVC, конечности E12.5 и общих сайтах связывания.
Анализ мотива De novo пиков SOX9 ChIP-Seq с помощью SeqPos (Liu et al., 2011) позволил получить мономер SOX и матрицу веса положения димера SOX (PWM) (рис. 1C), которая аналогична мотив SOX9 JASPAR. Известно, что при определении пола SOX9 связывается в виде мономера с одним ДНК-связывающим мотивом в регуляторных областях локусов стероидогенного фактора 1 (9).0003 Nr5a1 ) и антимюллеров гормон ( Amh ), тогда как гены, необходимые для хондрогенеза, такие как Col10a1 и Col9a1 , имеют два мотива связывания SOX9, разделенных четырьмя нуклеотидами в обратной и комплементарной ориентации друг другу, где SOX9 связывается как гомодимер (Bernard et al., 2003). Это аналогично мотиву димера SOX9, обнаруженному в наших данных. Чтобы определить, сколько пиков SOX9 содержат мотив мономера и/или димера SOX, мы просканировали все пики SOX9 с помощью Screen Motif (инструмент, написанный Клиффом Мейером и Леном Тайгом в Cistrome; Liu et al. , 2011), используя ШИМ, сгенерированные SeqPos. . Мы обнаружили, что 77% SOX9пики-конечности и 58% пиков SOX9 E12.5 AVC содержали по крайней мере один мономерный или димерный мотив (таблица S5). Во многих случаях пики SOX9 содержат несколько ДНК-связывающих мотивов SOX9 на пик. Последовательность димера SOX была идентифицирована в 34% пиков конечностей SOX9 по сравнению с 13,5% пиков SOX9 AVC, что позволяет предположить, что SOX9 в основном связывается в виде мономера с сердечными клапанами E12.5.
, 2011), используя ШИМ, сгенерированные SeqPos. . Мы обнаружили, что 77% SOX9пики-конечности и 58% пиков SOX9 E12.5 AVC содержали по крайней мере один мономерный или димерный мотив (таблица S5). Во многих случаях пики SOX9 содержат несколько ДНК-связывающих мотивов SOX9 на пик. Последовательность димера SOX была идентифицирована в 34% пиков конечностей SOX9 по сравнению с 13,5% пиков SOX9 AVC, что позволяет предположить, что SOX9 в основном связывается в виде мономера с сердечными клапанами E12.5.
 При взвешивании ассоциаций таким образом пики SOX9, локализованные во внутригенных регионах, были связаны с этими генами, а не с потенциально более близким TSS. Эта система картирования позволила нам связать пики с генами с меньшим количеством потенциальных ложных срабатываний и гарантировала, что большинство пиков были связаны только с одним геном. Примерно 22% и 31% SOX9пики в лимбе и AVC, соответственно, располагались либо непосредственно над TSS, либо в проксимальных областях промотора размером 5 kb. Это отличается от членов семейства SOX9, SOX6, у которых было только 13,5% сайтов связывания в TSS и 20 т.п.н. выше по течению (An et al., 2011), и SOX3, у которого было 11% сайтов связывания в проксимальных областях промотора. (Макэнинч и Томас, 2014 г.). Более того, заметные 63,6% пиков SOX9, общих для AVC и конечности (497 из 782), были расположены в промоторных областях TSS/5 kb (рис. 1D). Эти данные указывают на то, что SOX9часто связывается с промоутерами.
При взвешивании ассоциаций таким образом пики SOX9, локализованные во внутригенных регионах, были связаны с этими генами, а не с потенциально более близким TSS. Эта система картирования позволила нам связать пики с генами с меньшим количеством потенциальных ложных срабатываний и гарантировала, что большинство пиков были связаны только с одним геном. Примерно 22% и 31% SOX9пики в лимбе и AVC, соответственно, располагались либо непосредственно над TSS, либо в проксимальных областях промотора размером 5 kb. Это отличается от членов семейства SOX9, SOX6, у которых было только 13,5% сайтов связывания в TSS и 20 т.п.н. выше по течению (An et al., 2011), и SOX3, у которого было 11% сайтов связывания в проксимальных областях промотора. (Макэнинч и Томас, 2014 г.). Более того, заметные 63,6% пиков SOX9, общих для AVC и конечности (497 из 782), были расположены в промоторных областях TSS/5 kb (рис. 1D). Эти данные указывают на то, что SOX9часто связывается с промоутерами. Мы идентифицировали 2453 и 5750 генов со связанными пиками SOX9 в наборах данных E12. 5 AVC и ChIP-Seq конечностей соответственно (таблицы S3, S4). Примечательно, что 1605 генных локусов, которые имели ассоциированные пики SOX9, были обычными как в АВК, так и в конечностях. Из них 782 пика SOX9 перекрывались между двумя библиотеками, тогда как остальные локусы генов имели пики SOX9, связанные с разными геномными местоположениями в АВК и конечности (рис. 1B). Это указывает на то, что SOX9 взаимодействует с локусами генов, используя как общие, так и тканеспецифические регуляторные элементы.
5 AVC и ChIP-Seq конечностей соответственно (таблицы S3, S4). Примечательно, что 1605 генных локусов, которые имели ассоциированные пики SOX9, были обычными как в АВК, так и в конечностях. Из них 782 пика SOX9 перекрывались между двумя библиотеками, тогда как остальные локусы генов имели пики SOX9, связанные с разными геномными местоположениями в АВК и конечности (рис. 1B). Это указывает на то, что SOX9 взаимодействует с локусами генов, используя как общие, так и тканеспецифические регуляторные элементы.
Мы дополнительно определили общие функции SOX9, сравнив пики в AVC и конечностях с общедоступным набором данных SOX9 ChIP-Seq, который был создан в мышиных HF-SC (Kadaja et al., 2014) (рис. 2A). Мы идентифицировали 293 геномных местоположения с пиками SOX9 во всех трех библиотеках ChIP-Seq (рис. 2A; рис. S2; таблица S6), что позволяет предположить, что SOX9 занимает аналогичные области в мезенхимальных тканях и стволовых клетках. Анализ генной онтологии (GO) подмножества генов, связанных с этими общими областями, взаимодействующими с SOX9, с использованием категории биофункций (изобретательность) показал пролиферацию клеток (рис. 2B; таблица S7). Действительно, 106 генов, связанных с обычным SOX9пики были включены в категорию «пролиферация клеток» (таблица S8). Регуляторы клеточного цикла, связанные с пиками SOX9 во всех трех библиотеках, включают Junb , Cops5 , Fosl1 , Fosl2 и Fos . Кроме того, несколько генов, участвующих в пролиферации клеток, имели пики SOX9 в сердечном клапане и конечностях, но не в HF-SC (рис. 2C), например, Trp53 и Fgfr2 . ChIP-qPCR на E12.5 AVC и конечности дополнительно подтвердили данные SOX9 ChIP-Seq, продемонстрировав, что эти общие области были обогащены SOX9.занятость по сравнению с IgG (рис. 2C).
2B; таблица S7). Действительно, 106 генов, связанных с обычным SOX9пики были включены в категорию «пролиферация клеток» (таблица S8). Регуляторы клеточного цикла, связанные с пиками SOX9 во всех трех библиотеках, включают Junb , Cops5 , Fosl1 , Fosl2 и Fos . Кроме того, несколько генов, участвующих в пролиферации клеток, имели пики SOX9 в сердечном клапане и конечностях, но не в HF-SC (рис. 2C), например, Trp53 и Fgfr2 . ChIP-qPCR на E12.5 AVC и конечности дополнительно подтвердили данные SOX9 ChIP-Seq, продемонстрировав, что эти общие области были обогащены SOX9.занятость по сравнению с IgG (рис. 2C).
Рис. 2.
Просмотреть большой Загрузить слайд
Гены с ассоциированными общими пиками SOX9 среди развивающихся тканей свидетельствуют о взаимной роли в пролиферации. (A) Диаграмма Венна геномного расположения пиков SOX9 в E12.5 AVC, E12.5 конечностей и стволовых клетках волосяных фолликулов (HF-SCs). (B) Верхние восемь терминов, показывающие обогащение на основе значений P из анализа GO для генов, связанных с 293 перекрывающимися пиками SOX9, и генов, уникально связанных с каждой тканью. (C) КПЦР, показывающая кратное обогащение SOX9ChIP по сравнению с IgG (среднее значение и стандартное отклонение трех ChIP) на E12.5 AVC и конечности подтверждают пики SOX9, которые являются общими во всех трех библиотеках SOX9 ChIP-Seq и связаны с генами, участвующими в пролиферации клеток. Область, ранее идентифицированная в хондроцитах как SOX9-связанный энхансер Col2a1 , использовали в качестве положительного контроля, а неродственные геномные области использовали в качестве отрицательного контроля. (D) ChIP-qPCR показывает, что SOX9 связывается с одними и теми же областями в легком, кишечнике и печени E12.5 (три ChIP для Wasf1/2 , Hdac1/2 ; две микросхемы для Junb , p53 , Fgf11 , Col2a1 , Apoc , Tat ; и один чип для Fos и Eed ).
(B) Верхние восемь терминов, показывающие обогащение на основе значений P из анализа GO для генов, связанных с 293 перекрывающимися пиками SOX9, и генов, уникально связанных с каждой тканью. (C) КПЦР, показывающая кратное обогащение SOX9ChIP по сравнению с IgG (среднее значение и стандартное отклонение трех ChIP) на E12.5 AVC и конечности подтверждают пики SOX9, которые являются общими во всех трех библиотеках SOX9 ChIP-Seq и связаны с генами, участвующими в пролиферации клеток. Область, ранее идентифицированная в хондроцитах как SOX9-связанный энхансер Col2a1 , использовали в качестве положительного контроля, а неродственные геномные области использовали в качестве отрицательного контроля. (D) ChIP-qPCR показывает, что SOX9 связывается с одними и теми же областями в легком, кишечнике и печени E12.5 (три ChIP для Wasf1/2 , Hdac1/2 ; две микросхемы для Junb , p53 , Fgf11 , Col2a1 , Apoc , Tat ; и один чип для Fos и Eed ). Стандартное отклонение ChIP-qPCR показано в виде столбцов погрешностей на C и D. Отрицательные 1, 2, 3 ( Apoc3, Hnf1, Tat ) представляют собой области отрицательного контроля, которые не содержат пика SOX9. Регуляторная область Col2a1 представляет собой положительную контрольную область для связывания SOX9.
Стандартное отклонение ChIP-qPCR показано в виде столбцов погрешностей на C и D. Отрицательные 1, 2, 3 ( Apoc3, Hnf1, Tat ) представляют собой области отрицательного контроля, которые не содержат пика SOX9. Регуляторная область Col2a1 представляет собой положительную контрольную область для связывания SOX9.
Рис. 2.
Посмотреть большойСкачать слайд
Гены с ассоциированными общими пиками SOX9 среди развивающихся тканей свидетельствуют о взаимной роли в пролиферации. (A) Диаграмма Венна геномного расположения пиков SOX9 в E12.5 AVC, E12.5 конечностей и стволовых клетках волосяных фолликулов (HF-SCs). (B) Верхние восемь терминов, показывающие обогащение на основе значений P из анализа GO для генов, связанных с 293 перекрывающимися пиками SOX9, и генов, уникально связанных с каждой тканью. (C) КПЦР, показывающая кратное обогащение SOX9ChIP по сравнению с IgG (среднее значение и стандартное отклонение трех ChIP) на E12. 5 AVC и конечности подтверждают пики SOX9, которые являются общими во всех трех библиотеках SOX9 ChIP-Seq и связаны с генами, участвующими в пролиферации клеток. Область, ранее идентифицированная в хондроцитах как SOX9-связанный энхансер Col2a1 , использовали в качестве положительного контроля, а неродственные геномные области использовали в качестве отрицательного контроля. (D) ChIP-qPCR показывает, что SOX9 связывается с одними и теми же областями в легком, кишечнике и печени E12.5 (три ChIP для Wasf1/2 , Hdac1/2 ; две микросхемы для Junb , p53 , Fgf11 , Col2a1 , Apoc , Tat ; и один чип для Fos и Eed ). Стандартное отклонение ChIP-qPCR показано в виде столбцов погрешностей на C и D. Отрицательные 1, 2, 3 ( Apoc3, Hnf1, Tat ) представляют собой области отрицательного контроля, которые не содержат пика SOX9. Регуляторная область Col2a1 представляет собой положительную контрольную область для связывания SOX9.
5 AVC и конечности подтверждают пики SOX9, которые являются общими во всех трех библиотеках SOX9 ChIP-Seq и связаны с генами, участвующими в пролиферации клеток. Область, ранее идентифицированная в хондроцитах как SOX9-связанный энхансер Col2a1 , использовали в качестве положительного контроля, а неродственные геномные области использовали в качестве отрицательного контроля. (D) ChIP-qPCR показывает, что SOX9 связывается с одними и теми же областями в легком, кишечнике и печени E12.5 (три ChIP для Wasf1/2 , Hdac1/2 ; две микросхемы для Junb , p53 , Fgf11 , Col2a1 , Apoc , Tat ; и один чип для Fos и Eed ). Стандартное отклонение ChIP-qPCR показано в виде столбцов погрешностей на C и D. Отрицательные 1, 2, 3 ( Apoc3, Hnf1, Tat ) представляют собой области отрицательного контроля, которые не содержат пика SOX9. Регуляторная область Col2a1 представляет собой положительную контрольную область для связывания SOX9.
Учитывая, что SOX9 необходим для развития многих различных органов и, как известно, участвует в пролиферации клеток-предшественников (обзор Pritchett et al., 2011), мы ожидали, что общие пики SOX9, связанные с генами пролиферации, также будут присутствуют в других экспрессирующих SOX9 тканях. ChIP-qPCR была проведена на легком, кишечнике и печени E12.5 и продемонстрировала, что многие из общих пиковых областей SOX9 также были обогащены в этих тканях (рис. 2D). В совокупности наши данные свидетельствуют о том, что аналог SOX9ДНК-связывающие области используются в регуляции клеточной пролиферации как в различных типах клеток, так и во многих развивающихся тканях.
 2B). SOX9, специфичный для конечностей-ассоциированные гены выявили уникальные термины GO, участвующие в развитии мезенхимы, организации ECM и морфогенезе передних конечностей (Fig. 2B). SOX9 AVC-специфические гены-мишени идентифицировали уникальные категории GO, участвующие в связывании ДНК, развитии клеток сердечного нервного гребня (NCC) и морфогенезе восходящей аорты. Идентификация генов, вовлеченных в кардиальный NCC в AVC-специфических категориях GO, подтверждает сходство между AVC и OFT в развитии подушки. В целом, гены, связанные с тканеспецифичными пиками SOX9, сильно отражают уникальные характеристики каждой ткани, и это предполагает, что SOX9играет роль в тканевой идентичности и тесно связана с ней.
2B). SOX9, специфичный для конечностей-ассоциированные гены выявили уникальные термины GO, участвующие в развитии мезенхимы, организации ECM и морфогенезе передних конечностей (Fig. 2B). SOX9 AVC-специфические гены-мишени идентифицировали уникальные категории GO, участвующие в связывании ДНК, развитии клеток сердечного нервного гребня (NCC) и морфогенезе восходящей аорты. Идентификация генов, вовлеченных в кардиальный NCC в AVC-специфических категориях GO, подтверждает сходство между AVC и OFT в развитии подушки. В целом, гены, связанные с тканеспецифичными пиками SOX9, сильно отражают уникальные характеристики каждой ткани, и это предполагает, что SOX9играет роль в тканевой идентичности и тесно связана с ней. Чтобы проанализировать ключевые мишени транскрипции SOX9 во время развития сердечного клапана, мы воспользовались моделью мыши со специфической делецией Sox9 , в которой как эндокардиальные клетки, так и недавно трансформированные мезенхимальные клетки развивающихся подушек АВК не могут экспрессировать SOX9 (Lincoln и др. , 2007). Было показано, что эти мыши умирают во время эмбриогенеза и имеют дефекты сердечных клапанов, включая гипоплазию сердечных подушек, сниженную пролиферацию мезенхимы и измененный состав внеклеточного матрикса. 9Ранее было показано, что мышь 0003 Tie2-Cre , используемая для этой условной системы, специфически экспрессируется в эндокардиальных клетках и образующихся мезенхимальных клетках, а не в кардиомиоцитах, эпикарде или дистальной мезенхиме OFT (Kisanuki et al., 2001; Snarr et al., 2008). Чтобы гарантировать делецию Sox9 в E12.5 AVC, иммунофлуоресценцию SOX9 проводили на Sox9 fl/fl (дикий тип; WT) и Sox9 fl/fl ; cKO) сердца эмбрионов (рис. S3A,B; рис. S4). Как и ожидалось, удаление Sox9 приводит к гибели большинства эмбрионов на стадии E13.5 из-за сильно гипопластических и деформированных атриовентрикулярных клапанов (рис. S3A, B; рис. S4). Кроме того, у сердец Sox9 cKO стенки желудочков были истончены (рис.
, 2007). Было показано, что эти мыши умирают во время эмбриогенеза и имеют дефекты сердечных клапанов, включая гипоплазию сердечных подушек, сниженную пролиферацию мезенхимы и измененный состав внеклеточного матрикса. 9Ранее было показано, что мышь 0003 Tie2-Cre , используемая для этой условной системы, специфически экспрессируется в эндокардиальных клетках и образующихся мезенхимальных клетках, а не в кардиомиоцитах, эпикарде или дистальной мезенхиме OFT (Kisanuki et al., 2001; Snarr et al., 2008). Чтобы гарантировать делецию Sox9 в E12.5 AVC, иммунофлуоресценцию SOX9 проводили на Sox9 fl/fl (дикий тип; WT) и Sox9 fl/fl ; cKO) сердца эмбрионов (рис. S3A,B; рис. S4). Как и ожидалось, удаление Sox9 приводит к гибели большинства эмбрионов на стадии E13.5 из-за сильно гипопластических и деформированных атриовентрикулярных клапанов (рис. S3A, B; рис. S4). Кроме того, у сердец Sox9 cKO стенки желудочков были истончены (рис. S3A, звездочка), а межпредсердная и желудочковая перегородки не слились с АВК в Sox9 cKO по сравнению с WT (рис. S3A, стрелки). Иллюстрируя специфичность делеции Sox9 линии Tie2-Cre , экспрессия SOX9 была потеряна только в AVC, тогда как эпикардиальные клетки, происходящие из другой линии, все еще экспрессируют SOX9.(рис. S3A,B; рис. S4). Также было отмечено снижение пролиферации в культурах Sox9 cKO AVC и эксплантатов AVC (рис. S5), как видно из предыдущей работы (Lincoln et al., 2007).
S3A, звездочка), а межпредсердная и желудочковая перегородки не слились с АВК в Sox9 cKO по сравнению с WT (рис. S3A, стрелки). Иллюстрируя специфичность делеции Sox9 линии Tie2-Cre , экспрессия SOX9 была потеряна только в AVC, тогда как эпикардиальные клетки, происходящие из другой линии, все еще экспрессируют SOX9.(рис. S3A,B; рис. S4). Также было отмечено снижение пролиферации в культурах Sox9 cKO AVC и эксплантатов AVC (рис. S5), как видно из предыдущей работы (Lincoln et al., 2007).
Чтобы идентифицировать нижестоящие изменения мРНК, возникающие в результате делеции Sox9 , которые могут быть ответственны за клапанные дефекты, мы сравнили транскриптом E12.5 AVC у WT с транскриптомом Sox9 cKO. Поскольку эффективность делеции Sox9 в AVC с помощью Tie2-Cre является вариабельным, РНК была выделена из отдельных AVC из E12.5 WT и Sox9 cKO, а qRT-PCR подтвердила потерю транскрипта Sox9 перед объединением двух-трех AVC для библиотек RNA-Seq. Дублирующие библиотеки были созданы для каждого генотипа (рис. S6). При сравнении 634 гена были подавлены в Sox9 cKO как минимум в 1,5 раза, а 610 генов были активизированы в Sox9 cKO как минимум в 1,5 раза (таблицы S10, S11). Интересно, что подавление генов в Sox9 cKO AVC были классифицированы GO как участвующие в развитии хряща, дифференцировке мезенхимы и EMT (рис. 3A). Активированные гены в Sox9 cKO включали такие функции, как ответ на окислительный стресс, транспорт газа и положительную регуляцию сердечного ритма (рис. 3B).
Дублирующие библиотеки были созданы для каждого генотипа (рис. S6). При сравнении 634 гена были подавлены в Sox9 cKO как минимум в 1,5 раза, а 610 генов были активизированы в Sox9 cKO как минимум в 1,5 раза (таблицы S10, S11). Интересно, что подавление генов в Sox9 cKO AVC были классифицированы GO как участвующие в развитии хряща, дифференцировке мезенхимы и EMT (рис. 3A). Активированные гены в Sox9 cKO включали такие функции, как ответ на окислительный стресс, транспорт газа и положительную регуляцию сердечного ритма (рис. 3B).
Рис. 3.
Просмотреть большой Загрузить слайд
Сравнение SOX9 ChIP-Seq и дифференциальных транскриптов в Sox9 cKO AVC выявляет критические гены, участвующие в формировании клапана. (A,B) GO-анализ 634 генов с пониженной (A) и 610 повышающей (B) генов с ≥1,5-кратным изменением E12.5 Sox9 cKO. (C) Ingenuity идентифицирует сеть, включающую биофункции для транскрипции ДНК, кардиогенеза и аномальной морфологии сердца в генах с повышенной и пониженной регуляцией, которые имеют связанный пик SOX9. Линии указывают тип связи между геном и термином GO. Например, потеря Pitx2 приводит к ингибированию кардиогенеза и как к активации, так и к ингибированию транскрипции ДНК. Желтые линии указывают на наличие данных, подтверждающих как активацию, так и ингибирование. Библиотеки были созданы в двух экземплярах из объединенных клапанов.
Линии указывают тип связи между геном и термином GO. Например, потеря Pitx2 приводит к ингибированию кардиогенеза и как к активации, так и к ингибированию транскрипции ДНК. Желтые линии указывают на наличие данных, подтверждающих как активацию, так и ингибирование. Библиотеки были созданы в двух экземплярах из объединенных клапанов.
Рис. 3.
Просмотреть большой Загрузить слайд
Сравнение SOX9 ChIP-Seq и дифференциальных транскриптов в Sox9 cKO AVC выявляет критические гены, участвующие в формировании клапана. (A,B) GO-анализ 634 генов с пониженной (A) и 610 повышающей (B) генов с ≥1,5-кратным изменением E12.5 Sox9 cKO. (C) Ingenuity идентифицирует сеть, включающую биофункции для транскрипции ДНК, кардиогенеза и аномальной морфологии сердца в генах с повышенной и пониженной экспрессией, которые связаны с SOX9.вершина горы. Линии указывают тип связи между геном и термином GO. Например, потеря Pitx2 приводит к ингибированию кардиогенеза и как к активации, так и к ингибированию транскрипции ДНК. Желтые линии указывают на наличие данных, подтверждающих как активацию, так и ингибирование. Библиотеки были созданы в двух экземплярах из объединенных клапанов.
Желтые линии указывают на наличие данных, подтверждающих как активацию, так и ингибирование. Библиотеки были созданы в двух экземплярах из объединенных клапанов.
Для дальнейшего исследования генов, которые могут напрямую регулироваться SOX9, мы сосредоточились на 145 генах, экспрессия которых была изменена в AVC без Sox9 и имел связанный пик E12.5 AVC SOX9. Из этих 145 генов, для простоты называемых генами-мишенями SOX9, ~60% были подавлены, а 40% — активированы (таблицы 1, 2; таблица S12). Анализ генов-мишеней SOX9 с повышающей и понижающей регуляцией с помощью Ingenuity Pathway Analysis выявил обогащение функций, связанных с транскрипцией и кардиогенезом. Более того, некоторые из этих генов были связаны с аномальным морфогенезом сердца (Fig. 3C). Известные целевые TF, классифицированные как играющие роль в развитии сердца, которые были подавлены в Sox9 cKO включают Sox4 , Hand2 , Twist1 , Foxp4 , Mecom (также известный как Evi14 3) и 9000; гены-мишени с повышенной экспрессией также содержали несколько TF, таких как
Bhlhe40 , Ddit3 и Junb (таблицы 1, 2; таблица S12). Кроме того, несколько компонентов ВКМ, таких как периостин и эластин, были снижены (таблица 1; таблица S12). Таким образом, наши данные позволяют предположить, что SOX9 может действовать как активатор и репрессор транскрипции. Более того, наши данные подтверждают прямую роль SOX9.в регуляции сети TFs и компонентов ECM, которые функционируют во время развития сердечного клапана.
Table 1.
The top 20 direct targets of SOX9 with ≥1.5 fold change (FC) down in the Sox9 cKO AVC
View Large
Table 2.
The top 20 прямых мишеней SOX9 с ≥1,5-кратным изменением (FC) выше в Sox9 cKO AVC
View Large
3 и
(рис. 04,3 и 90). Во всех случаях экспрессия мРНК этих ТФ была существенно снижена в
Sox9 cKO AVCs. Два дополнительных TF, связанных с пиками SOX9, которые важны для развития сердечного клапана, Lef1 и Tbx20 , также были значительно снижены в Sox9 cKO AVC, как показано с помощью qRT-PCR (фиг. 4A), но были ниже 1,5- пороговое значение кратного изменения, используемое для анализа RNA-Seq ( Tbx20 = 1,33, Lef1 = 1,29; таблица S11). Следует отметить, что два наиболее подавленных гена ( Btn1a1 и Prelp ) и два наиболее активизированных гена ().0003 Bhlhle40 и Fos ) в Sox9 cKO AVC, также связанные с пиками SOX9, были подтверждены с помощью qRT-PCR (фиг. 4B). Интересно, что мало что известно о точной роли этих факторов в развитии сердечного клапана.Рис. 4.
Увеличить Загрузить слайд
SOX9 регулирует сеть ТФ во время развития клапана. (A) qRT-PCR для экспрессии мРНК Sox9 и выбранных целевых TF в E12.5 WT и Sox9 cKO AVC. ( n =8+ для Sox9 , Twist1 , Lef1 , Tbx20 ; n =3+ для Sox4 , Mecom , Pitx2 , Hand2 , Nfia ). Столбики погрешностей представляют собой sem. Относительную количественную оценку проводили с использованием Actb для qRT-PCR или с Gapdh для анализа Taqman в качестве контроля. (B) qRT-PCR на двух верхних генах с пониженной и повышенной экспрессией, которые также имеют связанные пики SOX9 ( n = 2). (C) Анализ люциферазы на HEK29Клетки 3T, связанные со сверхэкспрессией SOX9 или без нее, с использованием регуляторных областей, идентифицированных по пикам SOX9 ChIP-Seq для Mecom (энхансер), Nfia (энхансер) или Junb (промотор). n ≥3. (D) Анализ люциферазы с направленным мутагенезом для энхансера Mecom . Энхансерная область Mecom содержала как димерный, так и мономерный мотив SOX9, и сайты были мутированы либо только для димерного сайта (DM), либо для всех сайтов (allM). n ≥3. (Э) in situ Гибридизация для SOX9 и Twist1 на E12.5 WT ( SOX9 FL/FL ;+/+ ) и SOX9 CKO ( SOX9 FL/FL ; TIE2-CRE ( SOX9 FL/FL ; TIE2-CRE ( SOX9 FL/FL ; TIE2-CRE ( SOX9 FL/FL ; + ) сердца. AS, антисмысловой зонд, меченный дигоксигенином. * P <0,05, ** P <0,01, определено с помощью t -тестов по методу Сидака-Бонферрони. Столбики погрешностей представляют собой стандартное отклонение (sd), если не указано иное. Шкала баров: 200 мкм.
Рис. 4.
Увеличить Загрузить слайд
SOX9 регулирует сеть ТФ во время разработки клапана. (A) qRT-PCR для экспрессии мРНК Sox9 и выбранных целевых TF в E12.5 WT и Sox9 cKO AVC. ( N = 8+ для SOX9 , Twist1 , LEF1 , TBX20 ; N = 3+ для SOX4 , MECOM , PITX2 , , NFIA, 44444444444, 4444444, , , , 444444, , 444444, , , 4, , , 4, 4, , , , NFIA , . ). Столбики погрешностей представляют собой sem. Относительную количественную оценку проводили с Actb для qRT-PCR или с Gapdh для анализов Taqman в качестве контроля. (B) qRT-PCR на двух верхних генах с пониженной и повышенной экспрессией, которые также имеют связанные пики SOX9 ( n = 2). (C) Анализы люциферазы на клетках HEK293T в сочетании со сверхэкспрессией SOX9 или без нее с использованием регуляторных областей, идентифицированных пиками SOX9 ChIP-Seq для Mecom (энхансер), Nfia (энхансер) или Junb (промотор). n ≥3. (D) Анализы люциферазы с сайт-направленным мутагенезом для Усилитель Меком . Энхансерная область Mecom содержала как димерный, так и мономерный мотив SOX9, и сайты были мутированы либо только для димерного сайта (DM), либо для всех сайтов (allM). n ≥3. (E) in situ Гибридизация для SOX9 и Twist1 на E12.5 WT ( SOX9 FL/FL ;+/+ ) и SOX9 CKO ( SOX9 FL/FL 9237 -Cre/+ ) сердца. AS, антисмысловой зонд, меченный дигоксигенином. * Р <0,05, ** P <0,01, определено с помощью t -тестов по методу Сидака-Бонферрони. Столбики погрешностей представляют собой стандартное отклонение (sd), если не указано иное. Шкала баров: 200 мкм.
Для подтверждения того, что SOX9 может действовать как активатор и репрессор транскрипции, области пиков SOX9, связанные с Mecom , Nfia и Junb , были клонированы в репортерные векторы люциферазы и вместе с эктопической экспрессией SOX9 (pcDNA3-S ), активность люциферазы анализировали в HEK293T-клетки (рис. 4C). Пики SOX9, связанные с Mecom- и Nfia , располагались выше генов в предполагаемых энхансерах, тогда как пик SOX9 для Junb располагался внутри промотора. Было показано, что Mecom экспрессируется в развивающихся клапанах сердца (Hoyt et al., 1997; Bard-Chapeau et al., 2014), но его точная роль в атриовентрикулярных клапанах изучена недостаточно. Было показано, что факторы NFI работают вместе с SOX9 для регуляции нижестоящих генов-мишеней в других системах (Nagy et al., 2011; Kang et al., 2012) и Junb играет хорошо зарекомендовавшую себя роль в регуляции клеточного цикла. Эктопическая экспрессия SOX9 активировала энхансеры Mecom и Nfia в 2,45 и 2,37 раза соответственно, тогда как SOX9 ингибировала активность промотора Junb в 1,5 раза (фиг. 4C). Эти данные подтверждают предположение, что SOX9 может функционировать как для активации, так и для репрессии транскрипции.
Предполагаемый энхансер для Mecom содержит как димерный мотив SOX9, так и несколько мономерных мотивов. Чтобы показать, что эти SOX9мотивы необходимы для регуляции SOX9 этого энхансера, мы создали две мутантные версии: одну с мутированным димерным мотивом SOX9 (DM) и одну с мутированными димерными и мономерными мотивами (рис. S7). Делеция димерного мотива SOX9 привела к существенной потере активности SOX9-зависимой люциферазы из энхансера Mecom , а мутация всех потенциальных мотивов SOX9 привела к полной потере SOX9-зависимой активности (рис. 4D). Эти данные предполагают, что димерный мотив SOX9 в Mecom 9Энхансер 0004 необходим для большинства SOX9-зависимых индукций.
TWIST1 в высокой степени экспрессируется во время формирования мезенхимы сердечной подушки и играет важную роль в пролиферации и дифференцировке мезенхимы клапана (Shelton and Yutzey, 2008; Chakraborty et al., 2010b). Из-за ключевой роли TWIST1 в развитии сердечных клапанов мы сосредоточились на характеристике уровней транскриптов Twist1 в клапанах Sox9 cKO. qRT-PCR подтвердил, что 9Транскрипт 0003 Twist1 не только редуцирован на E12. 5, как обсуждалось, но также редуцирован в более ранний момент времени (E10.5) в Sox9 cKO AVC (рис. S8). Используя гибридизацию in situ , мы обнаружили, что мРНК Twist1 специфически экспрессируется в мезенхиме клапана в E12.5 AVC (рис. 4E; рис. S9). Хотя точный количественный анализ с помощью гибридизации in situ невозможен, мРНК Twist1 показала такой же уровень снижения, как и qRT-PCR в 9Мутантные клапаны 0003 Sox9 cKO на ст. E12.5 (рис. 4A). Несмотря на in vivo снижение экспрессии Twist1 в Sox9 -дефицитных AVC, энхансер, связанный с Twist1 , является высокоактивным без сверхэкспрессии SOX9 с использованием люциферазных анализов (данные не показаны). Анализ мотивов на Twist1 -ассоциированном энхансере идентифицировал центрально расположенный мотив NF-Y, а недавняя работа предполагает, что NF-Y может рекрутировать SOX9 (Shi et al., 2015).
Общая идентификация SOX9Пики ChIP-Seq в E12. 5 AVC и изменения уровней мРНК между WT и Sox9 cKO AVC подтверждают модель, в которой SOX9 функционирует как для активации, так и для подавления транскрипции во время развития сердечного клапана. SOX9 играет важную роль в регуляции специфичных для сердечных клапанов регуляторных сетей путем прямой активации факторов транскрипции, таких как Mecom и Nfia . Однако SOX9 может также косвенно регулировать такие факторы, как Twist1 , посредством взаимодействия с NF-Y или дополнительными партнерами по связыванию.
Во время развития сердечного клапана SOX9 сильно экспрессируется в мезенхиме кардиальных подушек АВК и OFT (Akiyama et al., 2004). Несмотря на важную роль SOX9 в развитии сердечного клапана, мало что известно о его механизмах или генах, регулируемых SOX9. Наше исследование показало двойную роль SOX9 в АВК за счет регуляции пролиферации клеток мезенхимы и за счет модулирования ключевых факторов транскрипции, которые функционируют во время развития сердечного клапана. На сегодняшний день наше исследование является единственным, в котором изучается SOX9.Геномные взаимодействия в эмбриональных тканях сердца и конечностей.
Хотя многие пики SOX9 не перекрываются, мы идентифицировали почти 300 контекстно-независимых пиков SOX9, которые расположены в аналогичных областях генома в трех различных исследованных тканях. Многие из этих общих пиков SOX9 связаны с генами, которые участвуют в пролиферации, что поддерживает общую функцию SOX9.в поддержании пролиферативного состояния во время эмбрионального развития (рис. 5). Давно известно, что SOX9 связан с пролиферацией. Однако прямая механистическая связь и транскрипционные мишени SOX9, участвующие в пролиферации, остаются неуловимыми. В AVC, конечностях и HF-SC пики SOX9 связаны с тремя членами семейства белков, трансформирующих Fos, Fos , Fosl1 и Fosl2 . Члены семейства FOS и JUN гетеродимеризуются с образованием комплекса AP-1, который, как известно, регулирует пролиферацию и выживание клеток, частично посредством экспрессии циклина D1 (Shaulian, 2010). В мезенхимальных стволовых клетках стабильный SOX9нокдаун вызывал снижение пролиферации, задержку прогрессирования S-фазы и повышение стабильности белка циклина D1 (Stockl et al., 2013). Следует отметить, что Junb также имеет связанный пик SOX9. Хотя JUNB сильно зависит от контекста, наиболее известно, что он ингибирует рост клеток, противодействуя активности JUN (Shaulian, 2010). Мы показываем, что экспрессия Fos и Junb повышается в SOX9-дефицитных AVC, предполагая, что их повышенная экспрессия может способствовать гипоплазии тканей.
Рис. 5.
Увеличить Загрузить слайд
Модель регуляции с помощью SOX9 сердечно-специфических и общих транскрипционных сетей. Черные стрелки указывают на взаимодействие, продемонстрированное нашими данными, а синие стрелки указывают на ранее опубликованные данные. Фиолетовые стрелки обозначают непрямые связи между молекулами. * Обратите внимание, что p53 нацелен на SOX9 только в AVC и конечности, а не в HF-SCs.
Рис. 5.
Посмотреть в большом размереСкачать слайд
Модель регулирования SOX9сердечно-специфических и общих транскрипционных сетей. Черные стрелки указывают на взаимодействие, продемонстрированное нашими данными, а синие стрелки указывают на ранее опубликованные данные. Фиолетовые стрелки обозначают непрямые связи между молекулами. * Обратите внимание, что p53 нацелен на SOX9 только в AVC и конечности, а не в HF-SCs.
В дополнение к факторам комплекса AP-1 несколько генов, играющих роль в клеточной пролиферации, имеют пики SOX9 в своих промоторах или потенциальных регуляторных областях, в том числе COPS5 , SRPK2 , AKT2 , EED , HDAC1 , HDAC2 , P53 ( TRP53 ) и PRKACA (PKA). COPS5 (сигналосомная субъединица 5 COP9) связывается с JUN-белками для повышения специфичности связывания и может разрушать ингибитор клеточного цикла p27Kip1 (Cdkn1b) (Claret et al., 1996). Потеря Cops5 в эмбриональной конечности приводит к укорочению конечностей из-за нарушения хондрогенеза, а уровни Sox9 снижаются в мутантных длинных костях (Bashur et al., 2014), что указывает на потенциальную петлю обратной связи между SOX9и COPS5 . SRPK2 (протеинкиназа 2 SRSF) может способствовать пролиферации и прогрессированию клеточного цикла за счет повышения уровня циклина D1 (Jang et al., 2009), а AKT2 регулирует прогрессирование клеточного цикла посредством фосфорилирования своих мишеней, включая ингибиторы циклинзависимой киназы и поддержание белка стабильность циклинов MYC и D-типа через GSK3β (Xu et al., 2012). EED (развитие эмбриональной эктодермы), HDAC1 и HDAC2 (гистондеацетилазы 1 и 2) являются эпигенетическими регуляторами, связанными с пролиферацией клеток (Bracken et al. , 2003; Kelly and Cowley, 2013). р53 активирует репарацию ДНК и останавливает пролиферацию, останавливая клеточный цикл. Если повреждение ДНК серьезное, p53 инициирует гибель клеток (Giono and Manfredi, 2006). PKA (протеинкиназа A) индуцируется циклическим аденозинмонофосфатом и регулирует клеточный рост и пролиферацию с помощью различных механизмов (Stork and Schmitt, 2002). Следует отметить, что PKA фосфорилирует SOX9.и увеличивает свою активность во время хондрогенеза (Huang et al., 2000). Активность p53 и PKA в клеточной пролиферации может быть специфичной для мезенхимы, поскольку пики SOX9 обнаруживаются в AVC и конечностях, но не в HF-SC.
В совокупности наши данные предполагают модель, в которой SOX9 способствует пролиферации во многих типах клеток во время развития, включая сердечные клапаны, путем взаимодействия с промоторами или энхансерами генов, кодирующих пролиферативные факторы, такие как белки AP-1, киназы и модификаторы гистонов. Поскольку многие из этих генов неправильно регулируются при различных заболеваниях, идентификация природы SOX9взаимодействия с регуляторными областями этих генов могут помочь выяснить, как контролируется регуляция этих генов.
Известно, что некоторые из этих генов-мишеней SOX9 важны для формирования клапана, включая гены, связанные с ECM, и TF. Пониженно регулируемые TF со связанными пиками SOX9 включали Lef1 , Pitx2 и Hand2 . Потеря Lef1 (из-за делеции TBX20), Pitx2 или Hand2 связана с дефектами клапанов (Liu et al. , 2002; Holler et al., 2010; Cai et al., 2013). ТФ с наиболее сниженной экспрессией в 9Клапаны 0003 Sox9 cKO были Twist1 , Sox4 и Mecom . Подобно SOX9, эти три TF сильно экспрессируются в подушках сердца, и мутации в каждом из этих факторов вызывают серьезные клапанные дефекты, приводящие к эмбриональной гибели (Ya et al., 1998; Vincentz et al., 2008; Bard-Chapeau et al. ., 2014).
Белки SOX, как известно, регулируют TF, которые будут функционировать как их будущие кофакторы (Kamachi and Kondoh, 2013). Известно, что SOX9 регулирует и взаимодействует с SOX5/6, чтобы регулировать гены-мишени в развивающейся конечности (Han and Lefebvre, 2008; Liu and Lefebvre, 2015), и оба они связаны с SOX9.пики в наших данных ChIP-Seq конечностей. Точно так же SOX9 может активировать SOX4 в сердце, чтобы помочь совместно регулировать гены, специфичные для клапана. Анализ мотивов выявил EVI1 (белковый продукт, полученный из Mecom ) как еще один потенциальный кофактор для SOX9, а сравнение пиков EVI1 в раковых клетках (Bard-Chapeau et al. , 2012) с пиками SOX9 AVC выявило сотни перекрывающихся генов-мишеней (Bard-Chapeau et al., 2012) P.A.H., VCG и R.C., неопубликованные).
Мы показали, что SOX9 может модулировать уровни Twist1 выражение в развивающемся AVC. В отсутствие SOX9 экспрессия мРНК Twist1 снижалась примерно в три раза в мезенхиме створки. TWIST1 может индуцировать пролиферацию и миграцию мезенхимы клапана во время раннего формирования клапана (Shelton and Yutzey, 2008; Chakraborty et al., 2010b), а после EMT TWIST1 играет роль в регуляции дифференцировки мезенхимы АВК (Vrljicak et al., 2012). ). Когда TWIST1 сохраняется на более поздних стадиях развития клапана, это приводит к повышенной пролиферации мезенхимы, повышенной экспрессии TBX20 и примитивному ECM (Chakraborty et al., 2010b). TWIST1 напрямую регулирует Tbx20 (Lee and Yutzey, 2011), но, следует отметить, Twist1 -null сердца не показывают различий в уровнях Tbx20 в АВК по сравнению с WT (Vincentz et al. , 2008). Интересно, что мы обнаружили, что TBX20 подавляется в Sox9 cKO AVC и что SOX9 занимает регуляторные области, связанные с TBX20. Эти данные предполагают, что TWIST1 и SOX9 могут сотрудничать, чтобы регулировать Tbx20 в развивающихся сердечных клапанах. Кроме того, SOX9 подавляется в Twist1- нулевой AVC (Vrljicak et al., 2012), что свидетельствует о существовании петли обратной связи между двумя факторами.
В совокупности наши данные показывают, что SOX9 занимает регуляторные области и влияет на экспрессию ключевых TF, которые жизненно важны для развития сердечного клапана (рис. 5). Интересно, что многие из этих TF регулируют экспрессию друг друга. Например, было показано, что TWIST1 регулирует Tbx20 (Lee and Yutzey, 2011), а TBX20, в свою очередь, регулирует LEF1 в клетках эндокарда клапана (Cai et al., 2013). Было показано, что EVI1 регулирует SOX4 и изменяет его экспрессию (Bard-Chapeau et al., 2014), а EVI1 и SOX4 могут сотрудничать вместе при миелоидном лейкозе (Boyd et al. , 2006). Это демонстрирует, что сложные взаимодействия происходят на нескольких уровнях регуляции транскрипции с помощью SOX9.и предполагает, что основные TF регулируются множеством способов, чтобы обеспечить правильное формирование клапана. Наша работа предполагает, что SOX9 и его транскрипционные мишени TF формируют генную регуляторную сеть, управляющую морфогенезом клапанов. У людей аберрантная экспрессия SOX9 и его транскрипционных мишеней связана с заболеванием клапанов сердца у взрослых (Hulin et al., 2013). Т.о., понимание SOX9-инициируемых сетей транскрипции в развитии сердечного клапана может дать дополнительную информацию о заболеваниях сердечного клапана у взрослых.
Мы хотели бы поблагодарить доктора Али Карсана за все его превосходные идеи и предложения в рамках этого проекта. Мы также хотели бы поблагодарить доктора Аманду Котцер за всю ее помощь в управлении этим проектом.
Авторские вклады
V.C.G., R.C., O.A. и П.А.Х. участвовал в постановке и планировании экспериментов. В.К.Г., Р.К., О.А. и Д.Ю.Л. проводил опыты. В.К.Г., Р.К., М.Б. и Р.В.В. проанализировал данные. В.К.Г., Р.К., Т.М.У. и П.А.Х. способствовал написанию рукописи. Ю.З., С.Дж.М.Дж. и М.А.М. участвовали в получении финансирования для проекта генома и/или в создании и обработке библиотек секвенирования.
Финансирование
Эта работа была поддержана грантом в помощь от Фонда сердца и инсульта Канады [G-14-0006139]; Геном Канады; и Геном Британской Колумбии.
Adam
,
R. C.
,
Yang
,
H.
,
Rockowitz
,
S.
,
Larsen
,
S. B.
,
Николова
,
М.
,
Oristian
,
D. S.
,
Polak
,
L.
,
Kadaja
,
M.
,
Asare
,
A.
,
Чжэн
,
Д.
и др. (
2015
).
Пионерские факторы управляют динамикой суперэнхансеров в отношении пластичности стволовых клеток и выбора линии
. .
https://doi.org/10.1038/nature14289
Akiyama
,
H.
,
Chaboissier
,
M.-C.
,
Martin
,
J. F.
,
Schedl
,
A.
4 и40002 де Кромбрюгге,
Б.
(
2002
).
Фактор транскрипции Sox9 играет важную роль на последовательных этапах пути дифференцировки хондроцитов и необходим для экспрессии Sox5 и Sox6
.
Гены Дев.
16
,
2813
—
2828
.
https://doi.org/10.1101/gad.1017802
Акияма
,
Х.
,
Шабуасье
,
М.-К.
,
Behringer
,
R. R.
,
Rowitch
,
D. H.
,
Schedl
,
A.
,
Epstein
,
J. A.
и
де Кромбрюгге
,
Б.
(
2004
).
Существенная роль Sox9 в пути, контролирующем образование сердечных клапанов и перегородок
.
Проц. Натл. акад. науч. США
101
,
6502
—
6507
.
https://doi.org/10.1073/pnas.0401711101
Ан
,
C.-I.
,
Донг
,
Ю.
и
Хагивара
,
Н.
(
2011
).
Полногеномное картирование сайтов связывания Sox6 в скелетных мышцах выявило как прямую, так и непрямую регуляцию терминальной дифференцировки мышц с помощью Sox6
.
BMC Dev. биол.
11
,
59
.
https://doi.org/10.1186/1471-213X-11-59
Бейкер
,
A.
,
Saltik
,
M.
,
Lehrmann
,
H.
,
Killisch
,
I.
,
Mautner
,
V.
,
Lamm
,
G.
,
Christofori
,
G.
and
Cotten
,
М.
(
1997
).
Полиэтиленимин (PEI) представляет собой простой, недорогой и эффективный реагент для конденсации и связывания плазмидной ДНК с аденовирусом для доставки генов
.
Джин Тер.
4
,
773
—
782
.
https://doi.org/10.1038/sj.gt.3300471
Bard-Chapeau
,
E. A.
,
Jeyakani
,
J.
,
Kok
,
C. H.
,
Muller
,
J.
,
Chua
,
B. Q.
,
Gunaratne
,
J.
,
Batagov
,
A.
,
Jenjaroenpun
,
P.
,
Кузнецов
,
В. А.
,
Вэй
,
С. -Л.
и др. (
2012
).
Экотопический сайт интеграции вируса 1 (EVI1) регулирует множественные клеточные процессы, важные для рака, и является синергетическим партнером для белка FOS в инвазивных опухолях
.
Проц. Натл. акад. науч. США
109
,
2168
—
2173
.
https://doi.org/10.1073/pnas.11109
Bard-Chapeau
,
E. A.
,
Szumska
,
D.
,
Jacob
,
B.
,
Chua
,
B. Q. L.
,
Chatterjee
,
G. C.
,
Zhang
,
,
,
,
,
,
,
,
G. C.0002 Y.
,
Ward
,
J. M.
,
Urun
,
F.
,
Kinameri
,
E.
,
Vincent
,
S. D.
и др. (
2014
).
Мыши, несущие гипоморфный аллель Evi1, являются эмбрионально жизнеспособными, но имеют тяжелые врожденные пороки сердца
.
ПЛОС ОДИН
9
,
e89397
.
https://doi.org/10.1371/journal.pone.0089397
Bashur
,
L. A.
,
Chen
,
D.
,
Chen
,
Z.
,
Лян
,
Б.
,
Парди
,
Р.
Мураками
,
С.
и
Чжоу
,
Г.
(
2014
5).
Потеря jab1 в остеохондральных клетках-предшественниках серьезно нарушает развитие эмбриональных конечностей у мышей
.
Дж. Сотовый. Физиол.
229
,
1607
—
1617
.
https://doi.org/10.1002/jcp.24602
Bell
,
D. M.
,
Leung
,
K. K.
,
Wheatley
,
S. C.
,
Ng
,
L. J.
,
Zhou
,
S.
,
Wing Ling
,
K.
,
Har Sham
,
M.
,
Koopman
,
P.
,
TAM
,
P. P. L.
и
Cheah
,
K. S. E.
(
1997
000 000 000 000 000 000 000 000 000 9000.
SOX9 напрямую регулирует ген коллагена II типа
.
Нац. Жене.
16
,
174
—
178
.
https://doi.org/10.1038/ng0697-174
Bennebane
,
H.
и
Wrana
,
J. L.
(
2003
).
GATA- и Smad1-зависимые энхансеры в гене Smad7 по-разному интерпретируют концентрации костного морфогенетического белка
.
Мол. Клетка. биол.
23
,
6646
—
6661
.
https://doi. org/10.1128/MCB.23.18.6646-6661.2003
Bernard
,
P.
,
Tang
,
P.
,
LIU
,
S.
,
,
S.
,
,
,
,
4.44444444444444444.,
Harley
,
V. R.
и
Vilain
,
E.
(
2003
).
Димеризация SOX9 необходима для хондрогенеза, но не для определения пола
.
Гул. Мол. Жене.
12
,
1755
—
1765
.
https://doi.org/10.1093/hmg/ddg182
Blankenberg
,
D.
,
Von Kuster
,
G.
,
,
G.
,
,
G.
,
,
G.
.
,
Ананда
,
Г.
,
Lazarus
,
R.
,
Mangan
,
M.
,
Nekrutenko
,
A.
and
Taylor
,
J.
(
2010
).
Galaxy: веб-инструмент для анализа генома для экспериментаторов
.
Курс. протокол Мол. биол.
89
,
19.10.1
—
19.10.21
.
https://doi.org/10.1002/0471142727. mb1910s89
Бойд
,
К. Э.
,
Сяо
,
Y.-.
,
Fan
,
K.
,
Poholek
,
A.
,
Copeland
,
N. G.
,
Jenkins
,
Н. А.
и
Perkins
,
А. С.
(
2006 906).
Sox4 взаимодействует с Evi1 в миелоидных опухолях AKXD-23 посредством трансактивации провирусного LTR
.
Кровь
107
,
733
—
741
.
https://doi.org/10.1182/blood-2003-05-1626
Бракен
,
A. P.
,
Pasini
,
D.
,
Capra
,
M.
,
Prosperini
,
E.
,
Колли
,
Е.
и
Хелин
,
К.
(
2003
).
EZh3 находится ниже пути пути pRB-E2F, необходим для пролиферации и амплифицируется при раке
.
EMBO J.
22
,
5323
—
5335
.
https://doi.org/10.1093/emboj/cdg542
Bridgewater
,
L. C.
,
Lefebvre
,
V.
and
de Crombrugghe
,
B.
(
1998
).
Специфичные для хондроцитов энхансерные элементы в гене Col11a2 напоминают тканеспецифический энхансер Col2a1
.
Журнал биол. хим.
273
,
14998
—
15006
.
https://doi.org/10.1074/jbc.273.24.14998
CAI
,
X.
,
Zhang
,
W.
,
,
W.
,
,
W.
,
,
W.
,
,
W.
,
,
W.
, 9000 2,2 9000,
.
,
Чжан
,
Л.
,
Sultana
,
N.
,
WU
,
B.
,
CAI
,
W.
,
,
W.
,
,
W.
,
,
W.
,
,
W.
,
. и
Cai
,
C.-L.
(
2013
).
Tbx20 действует выше передачи сигналов Wnt, чтобы регулировать формирование эндокардиальной подушки и ремоделирование клапана во время кардиогенеза мыши
. .
https://doi.org/10.1242/dev.0
Chakraborty
,
S.
,
Combs
,
M. D.
and
Yutzey
,
K. E.
(
2010a
).
Транскрипционная регуляция клеток-предшественников сердечного клапана
.
Педиатр. Кардиол.
31
,
414
—
421
.
https://doi.org/10.1007/s00246-009-9616-x
Chakraborty
,
S.
,
Wirrig
,
E. E.
,
Hinton
,
Р. Б.
,
Меррилл
,
Ш. В.
,
Spicer
,
D. B.
и
Yutzey
,
K. E.
(
2010b
).
Twist1 способствует пролиферации клеток сердечного клапана и экспрессии генов внеклеточного матрикса во время развития in vivo и экспрессируется в пораженных аортальных клапанах человека
.
Дев. биол.
347
,
167
—
179
.
https://doi.org/10.1016/j.ydbio.2010.08.021
Chang
,
A. C. Y.
,
Fu
,
Y.
,
Garside
,
V. C.
,
Niessen
,
K.
,
Chang
,
L.
,
Fuller
,
9000. M.,
9000. M. M. M. M. M. M. M. M. M. M.999959 90009 9000 9000,
9000. M. 9000 9000 9000 9000 9000 9000,
9000. 9000.959 9000 9000 90000614 ,
Setiadi
,
A.
,
Smrz
,
J.
,
Kyle
,
A.
,
Minchinton
,
A
и др. (
2011
).
Notch инициирует эндотелиально-мезенхимальный переход в атриовентрикулярном канале посредством аутокринной активации растворимой гуанилатциклазы
.
Дев. Ячейка
21
,
288
—
300
.
https://doi.org/10.1016/j.devcel.2011.06.022
Chang
,
A. C. Y.
,
Garside
,
V. C.
,
Fournier
,
M
,
Смрз
,
Дж.
,
Vrljicak
,
P.
,
Umlandt
,
P.
,
Fuller
,
M.
,
Robertson
,
G.
,
Чжао
,
Ю.
,
Там
,
А.
, и др. (
2014
).
Notch-зависимая иерархия транскрипции способствует мезенхимальной трансдифференцировке в кардиальной подушке
.
Дев. Дин.
243
,
894
—
905
.
https://doi.org/10.1002/dvdy.24127
Бордовый
,
Ф.-Х.
,
Hibi
,
M.
,
Dhut
,
S.
,
Toda
,
T.
and
Karin
,
М.
(
1996
).
Новая группа консервативных коактиваторов, повышающих специфичность транскрипционных факторов АР-1
. .
https://doi.org/10.1038/383453a0
de Vlaming
,
A.
,
Sauls
,
K.
,
Hajdu
,
Z.
,
Visconti
,
R. P.
,
Mehesz
,
A. N.
,
Levine
,
R. A.
,
Slaugenhaupt
,
S. A.
,
Hagège
,
A.
,
Chester
,
А. Х.
,
Марквальд
,
Р. Р.
и др. (
2012
).
Развитие атриовентрикулярного клапана: новый взгляд на старую тему
. .
https://doi.org/10.1016/j.diff.2012.04.001
Эдем
,
E.
,
Navon
,
R.
,
Steinfeld
,
I.
,
Lipson
,
D.
and
Yakhini
,
З.
(
2009
).
GOrilla: инструмент для обнаружения и визуализации расширенных терминов GO в ранжированных списках генов
.
Биоинформатика BMC
10
,
48
.
https://doi.org/10.1186/1471-2105-10-48
Fejes
,
A. P.
,
Robertson
,
G.
,
Bilenky
,
М.
,
Вархол
,
Р.
,
Бейнбридж
,
М. и
05050002 Джонс,
С. Дж.
(
2008
).
FindPeaks 3.1: инструмент для выявления областей обогащения с помощью технологии массового параллельного секвенирования с коротким считыванием
.
Биоинформатика
24
,
1729
—
1730
https://doi.org/10.1093/bioinformatics/btn305
Джионо
,
L. E.
и
Manfredi
,
J. J.
(
2006
).
Опухолевой супрессор p53 участвует в нескольких контрольных точках клеточного цикла
.
J. Cell Physiol.
209
,
13
—
20
.
https://doi.org/10.1002/jcp.20689
Хан
,
Y.
и
Лефевр
,
В.
(
2008
).
L-Sox5 и Sox6 управляют экспрессией гена аггрекана в хрящах, обеспечивая связывание Sox9 с энхансером далеко выше по течению
.
Мол. Клетка. биол.
28
,
4999
—
5013
.
https://doi.org/10.1128/MCB.00695-08
Холлер
,
K. L.
,
Hendershot
,
T. J.
,
Troy
,
S. E.
,
Vincentz
,
J. W.
,
Firulli
,
A.B.
и
Howard
,
M. J.
(
2010
).
Направленная делеция Hand2 в клетках, происходящих из сердечного нервного гребня, влияет на экспрессию кардиального гена и развитие пути оттока
.
Дев. биол.
341
,
291
—
304
.
https://doi.org/10.1016/j.ydbio.2010.02.001
HOU
,
J.
,
Charters
,
A. M.
,
,
A. M.
,
,
A. M.
,
,
A. M.
4,,
A. M.
4,,
A. M.
44,
A.M. Ю. С.
,
Чжао
,
Ю.
,
Wu
,
M. K.
,
Jones
,
S. J. M.
,
Marra
,
M. A.
and
Hoodless
,
P. A.
(
2007
).
Систематический скрининг генов, экспрессируемых в дефинитивной энтодерме, с помощью серийного анализа экспрессии генов (SAGE)
.
BMC Dev. биол.
7
,
92
.
https://doi.org/10.1186/1471-213X-7-92
Hoyt
,
P. R.
,
Bartholomew
,
C.
,
Davis
,
А. Дж.
,
Ютзи
,
К.
,
Геймер
,
Л. В. 06040 9
Л. В. 06040
,
S. S.
,
IHLE
,
J. N.
и
Mucenski
,
M. L.
(
M. L.
(
M. L.
Протоонкоген Зла необходим в середине беременности для развития нервной системы, сердца и параксиальной мезенхимы
.
Мех. Дев.
65
,
55
—
70
.
https://doi.org/10.1016/S0925-4773(97)00057-9
Huang
,
W.
,
,
,
,
,
,
,
. ,
В.
и
де Кромбрюгге
,
Б.
(
2000
).
Фосфорилирование SOX9 циклической АМФ-зависимой протеинкиназой А усиливает способность SOX9 трансактивировать энхансер, специфичный для хондроцитов Col2a1
.
Мол. Клетка. биол.
20
,
4149
—
4158
.
https://doi.org/10.1128/MCB.20.11.4149-4158.2000
Hulin
,
A.
,
Deroanne
,
C.
,
Lambert
,
К.
,
Дефрейн
,
Ж.-О.
,
Nusgens
,
B.
,
Radermecker
,
M.
and
Colige
,
A.
(
2013
).
Новые патогенные механизмы миксоматозного митрального клапана человека: уроки прошлых и новых данных
.
Сердечно-сосудистые заболевания. Патол.
22
,
245
—
250
.
https://doi.org/10.1016/j.carpath.2012.11.001
Jain
,
V.
и
SEN
,
B.
(
,
B.
.
Кампомелическая дисплазия
.
J. Pediatr. Ортоп. Б
23
,
485
—
488
.
https://doi.org/10.1097/BPB.0000000000000058
Jang
,
S.-W.
,
Liu
,
X.
,
Fu
,
H.
,
Rees
,
H.
,
Yepes
,
М.
,
Леви
,
А.
и
Е
,
К.
5 ( 20 0 92055 ( 90 00055)).
Взаимодействие Akt-фосфорилированного SRPK2 с 14-3-3 опосредует клеточный цикл и гибель клеток в нейронах
.
Журнал биол. хим.
284
,
24512
—
24525
.
https://doi.org/10.1074/jbc.M109.026237
Кадая
,
М.
,
Кейс
, 09005 B.10614 , 09005 6 E
0005
Lin
,
M.
,
Pasolli
,
H. A.
,
Genander
,
M.
,
Polak
,
L.
,
Stokes
,
N.
,
Zheng
,
D.
и
Fuchs
,
E.
(
,
.(
,
.
(
,
.
4 (,
.
4 (,
9000 9000 9000 9000 9000 9000 9000 9000 9000..0614).
SOX9: регулятор транскрипции стволовых клеток секретируемых сигнальных факторов ниши
.
Гены Дев.
28
,
328
—
341
.
https://doi.org/10.1101/gad.233247.113
Kamachi
,
Y.
and
Kondoh
,
H.
(
2013
).
Белки Sox: регуляторы спецификации и дифференцировки клеточных судеб
. .
https://doi.org/10.1242/dev.0
Kang
,
P.
,
Lee
,
H. K.
,
Glasgow
,
S. M.
,
Finley
,
M.
,
Donti
,
T.
,
Gaber
,
Z. B.
,
Graham
,
B. H.
,
Foster
,
A. E.
,
Novitch
,
B. G.
,
gronostajski
,
.. M.M.
....4. 9000. 9000. 9000. 9000. 9000. 9000. 9000.. 9000. 9000. 9000. 9000. 9000. 9000. 9000. 9000. 9000. 9000. 9000.. (2012
).
Sox9 и NFIA координируют каскад регуляции транскрипции во время инициации глиогенеза
.
Нейрон
74
,
79
—
94
.
https://doi.org/10.1016/j.neuron.2012.01.024
Kelly
,
R.D.W.
и
Cowley
,
5 S. M.
0005(
2013
).
Физиологическая роль гистондеацетилазы (HDAC) 1 и 2: сложные партнеры с несколькими ведущими частями
.
Биохим. соц. Транс.
41
,
741
—
749
.
https://doi.org/10.1042/BST20130010
Кент
,
WJ
,
Сугнет
,
C. W.
,
Furey
,
T. S.
,
Roskin
,
K. M.
,
Pringle
,
T. H.
,
Zahler
,
AM
и
Haussler
,
D.
(
2002
).
Браузер генома человека в UCSC
.
Геном рез.
12
,
996
—
1006
.
https://doi.org/10.1101/gr.229102
Kim
,
D.
,
Pertea
,
G.
,
Trapnell
,
C.
,
Пиментел
,
Х.
,
Келли
,
Р.
и
Зальцберг
,
S. L.
(
2013
).
TopHat2: точное выравнивание транскриптомов при наличии вставок, делеций и слияний генов
.
Геном Биол.
14
,
R36
.
https://doi. org/10.1186/gb-2013-14-4-r36
Кисануки
,
YY
,
Молоток
,
Р. Е.
,
Миядзаки
,
J.-i.
,
Williams
,
S. C.
,
Richardson
,
J. A.
and
Yanagisawa
,
M.
(
2001
).
Трансгенные мыши Tie2-Cre: новая модель для анализа эндотелиальных клеточных линий in vivo
.
Дев. биол.
230
,
230
—
242
.
https://doi.org/10.1006/dbio.2000.0106
Lee
,
M. P.
and
Yutzey
,
K. E.
(
2011
).
Twist1 напрямую регулирует гены, которые способствуют пролиферации и миграции клеток в развивающихся сердечных клапанах
.
PLoS ONE
6
,
e29758
.
https://doi.org/10.1371/journal.pone.0029758
Lefebvre
,
V.
,
Huang
,
W.
,
Harley
,
V. R.
,
Goodfellow
,
PN
и
де Кромбрюгге
,
Б.
(
1997
).
SOX9 является мощным активатором специфического для хондроцитов энхансера гена коллагена pro alpha1(II)
.
Мол. Клетка. биол.
17
,
2336
—
2346
.
https://doi.org/10.1128/MCB.17.4.2336
Li
,
H.
и
Дурбин
,
Р.
(
2009
).
Быстрое и точное выравнивание коротких считываний с помощью преобразования Берроуза-Уилера
.
Биоинформатика.
https://doi.org/10.1093/bioinformatics/btp324
Lincoln
,
J.
,
Lange
,
А.В.
и
Ютзи
,
К.Е.
(
2006
5).
Сердце и кости: общие регуляторные механизмы развития сердечного клапана, хрящей, сухожилий и костей
.
Дев. биол.
294
,
292
—
302
.
https://doi. org/10.1016/j.ydbio.2006.03.027
Lincoln
,
J.
,
Kist
,
R.
,
Scherer
,
G.
and
Yutzey
,
K. E.
(
2007
).
Sox9 необходим для экспансии клеток-предшественников и организации внеклеточного матрикса во время развития сердечного клапана мыши
.
Дев. биол.
305
,
120
—
132
.
https://doi. org/10.1016/j.ydbio.2007.02.002
Лю
,
C.-F.
и
Лефевр
,
В.
(
2015
).
Факторы транскрипции SOX9 и SOX5/SOX6 взаимодействуют по всему геному посредством суперэнхансеров, чтобы управлять хондрогенезом
.
Рез. нуклеиновых кислот.
43
,
8183
—
8203
.
https://doi.org/10.1093/nar/gkv688
Liu
,
C.
,
Liu
,
W.
,
Palie
,
J.
,
Лу
,
М. Ф.
,
Коричневый
,
Н. А.
и
Martin
,
J. F.
(
2002
).
Pitx2c моделирует передние сосуды миокарда и дуги аорты и необходим для локального перемещения клеток в атриовентрикулярные подушки
. .
Лю
,
Т.
,
Ortiz
,
J. A.
,
Taing
,
L.
,
Meyer
,
C. A.
,
,
C. A.
,
,
C. A.
9000,,
C. A.
9000,,
C. A.
9000,,
C. A.
9000,,
с.
Zhang
,
Y.
,
Shin
,
H.
,
Wong
,
S.
,
,
S.
,
,
.0005
J.
,
Lei
,
Y.
, и др. (
2011
).
Cistrome: интегративная платформа для исследований регуляции транскрипции
.
Геном Биол.
12
,
R83
.
https://doi.org/10.1186/gb-2011-12-8-r83
McAninch
,
D.
и
Томас
,
П.
(
2014
).
Идентификация высококонсервативных предполагаемых энхансеров развития, связанных SOX3 в нейральных предшественниках, с использованием ChIP-Seq
.
PLoS ONE
9
,
e113361
.
https://doi.org/10.1371/journal.pone.0113361
Монтеро
,
Дж. А.
,
Giron
,
B.
,
Arrechedera
,
H.
,
Cheng
,
Y.-C.
,
Scotting
,
P.
,
Chimal-Monroy
,
J.
,
Garcia-Porrero
,
J. A.
and
Hurle
,
Дж. М.
(
2002
).
Экспрессия Sox8, Sox9 и Sox10 в развивающихся клапанах и вегетативных нервах эмбрионального сердца
.
Мех. Дев.
118
,
199
—
202
.
https://doi.org/10.1016/S0925-4773(02)00249-6
Надь
,
А.
,
Кенеси
4 ,
0 Э.0614 ,
Rentsendorj
,
O.
,
Molnar
,
A.
,
Szenasi
,
T.
,
Sinko
,
I .
,
Zvara
,
A.
,
Oommen
,
S. T.
,
Barta
,
E.
,
Puskas
,
Л. Г.
и др. (
2011
).
Эволюционно консервативная, специфичная для зоны роста регуляция промотора матрилина-1: факторы L-Sox5/Sox6 и Nfi, связанные рядом с ТАТА, тонко настраивают активацию с помощью Sox9
.
Мол. Клетка. биол.
31
,
686
—
699
.
https://doi.org/10.1128/MCB.00019-10
Ohba
,
S.
,
He
,
X.
,
Hojo
,
H.
and
McMahon
,
A. P.
(
2015
).
Различные программы транскрипции лежат в основе регуляции Sox9 хондроцитов млекопитающих
.
Сотовый представитель
12
,
229
—
243
.
https://doi. org/10.1016/j.celrep.2015.06.013
Person
,
A. D.
,
Klewer
,
S. E.
and
Runyan
,
R. B.
(
2005
).
Клеточная биология развития сердечной подушки
.
Междунар. Преподобный Цитол.
243
,
287
—
335
.
https://doi.org/10.1016/S0074-7696(05)43005-3
Pritchett
,
J.
,
Athwal
,
V.
,
Roberts
,
Н.
,
Hanley
,
Н.Д.0005
(
2011
).
Понимание роли SOX9 в приобретенных заболеваниях: уроки развития
.
Тенденции Мол. Мед.
17
,
166
—
174
.
https://doi.org/10.1016/j.molmed.2010.12.001
Робертсон
,
А.Г.
,
Беленький
,
5 М.
05
,
Tam
,
A.
,
Zhao
,
Y.
,
Zeng
,
T.
,
Thiessen
,
N.
,
Cezard
,
T.
,
Fejes
,
A. P.
,
Wederell
,
E. D.
,
Каллум
,
Р.
и др. (
2008
).
Полногеномная взаимосвязь между моно- и триметилированием лизина 4 гистона h4 и связыванием фактора транскрипции
.
Геном рез.
18
,
1906
—
1917
.
https://doi.org/10.1101/gr.078519.108
Робинсон
,
M. D.
,
McCarthy
,
D. J.
и
Smyth
,
G. K.
(
2010
4).edgeR: пакет Bioconductor для анализа дифференциальной экспрессии цифровых данных экспрессии генов
.
Биоинформатика
26
,
139
—
140
.
https://doi.org/10.1093/bioinformatics/btp616
Rockich
,
B. E.
,
Hrycaj
,
S. M.
,
Shih
,
H. P.
,
Nagy
,
M. S.
,
Ferguson
,
M. A. H.
,
KOPP
,
J. L.
,
Sander
,
M.
Sander
,
M.
,
M.
.0005
,
Wellik
,
D. M.
и
Spence
,
J. R.
(
2013
).
Sox9 играет несколько ролей в легочном эпителии во время морфогенеза ветвления
.
Проц. Натл. акад. науч. США
110
,
E4456
—
E4464
.
https://doi.org/10.1073/pnas.1311847110
Sekiya
,
I.
,
Tsuji
,
K.
,
Koopman
,
P.
,
Watanabe
,
H.
,
Yamada
,
Y.
,
Shinomiya
,
K.
,
Nifuji
,
A.
and
Noda
,
М.
(
2000
).
SOX9 усиливает активность промотора/энхансера гена аггрекана и активируется ретиноевой кислотой в клеточной линии, полученной из хряща, TC6
.
Журнал биол. хим.
275
,
10738
—
10744
.
https://doi. org/10.1074/jbc.275.15.10738
Шаулян
,
Э.
(
2010
).
AP-1 — Белки Jun: онкогены или замаскированные опухолевые супрессоры?
Сотовый. Сигнал.
22
,
894
—
899
.
https://doi.org/10.1016/j.cellsig.2009.12.008
Шелтон
,
Э. Л.
и
Ютзи
,
5 К. Э.0005
(
2008
).
Функция Twist1 при пролиферации, миграции и дифференцировке клеток эндокарда в процессе развития сердечного клапана
.
Дев. биол.
317
,
282
—
295
.
https://doi.org/10.1016/j.ydbio.2008.02.037
Ши
,
Z.
,
Чан
,
К.-И.
,
Labhart
,
P.
,
Zhao
,
Y.
,
Yang
,
J.
,
Mistretta
,
Т.-А.
,
Henning
,
S. J.
,
Maity
,
S. N.
и
Mori-Akiyama
,
и
Mori-Akiyama
,
и
Mori-Akiyama
,
и
Mori-Akiyama
,
.0005
Г.
(
2015
).
Контекстно-специфическая роль SOX9 в опосредованной NF-Y регуляции генов в клетках колоректального рака
.
Рез. нуклеиновых кислот.
43
,
6257
—
6269
.
https://doi.org/10.1093/nar/gkv568
Снарр
,
Б. С.
,
Керн
,
C.B.
и
Wessels
,
A.
(
2008
).
Происхождение и судьба сердечной мезенхимы
.
Дев. Дин.
237
,
2804
—
2819
.
https://doi.org/10.1002/dvdy.21725
Стокл
,
С.
,
Бауэр
,
R. J.
,
Bosserhoff
,
A. K.
,
Gottl
,
C.
,
Grifka
,
J.
and
Grassel
,
С.
(
2013
).
Sox9 модулирует выживаемость клеток и адипогенную дифференцировку мультипотентных мезенхимальных стволовых клеток взрослых крыс
.
J. Cell Sci.
126
,
2890
—
2902
.
https://doi. org/10.1242/jcs.124305
Стол
,
P. J. S.
и
Schmitt
,
J. M.
(
,
.
Перекрестные помехи между передачей сигналов цАМФ и МАР-киназы в регуляции клеточной пролиферации
.
Trends Cell Biol.
12
,
258
—
266
.
https://doi.org/10.1016/S0962-8924(02)02294-8
Timmerman
,
L. A.
,
Grego-Bessa
,
J.
,
Raya
,
А.
,
Бертран
,
Э.
,
Pérez-Pomares
,
J. M.
,
Díez
,
J.
,
Aranda
,
S.
,
Palomo
,
S.
,
McCormick
,
F.
,
Изписуа-Бельмонте
,
90 90 10 5 10 C. (2004
).
Notch способствует эпителиально-мезенхимальному переходу во время развития сердца и онкогенной трансформации
.
Гены Дев.
18
,
99
—
115
.
https://doi.org/10.1101/gad.276304
Trapnell
,
C.
,
Williams
,
B. A.
,
Pertea
,
G.
,
Мортазави
,
А.
,
Kwan
,
G.
,
van Baren
,
M. J.
,
Salzberg
,
S. L.
,
Wold
,
B. J.
and
Пачтер
,
Л.
(
2010
).
Сборка транскриптов и количественная оценка с помощью RNA-Seq выявляют неаннотированные транскрипты и переключение изоформ во время дифференцировки клеток
.
Нац. Биотехнолог.
28
,
511
—
515
.
https://doi.org/10.1038/nbt.1621
Trowe
,
М.-О.
,
Shah
,
S.
,
Petry
,
M.
,
Airik
,
R.
,
.0002 Schuster-Gossler
,
K.
,
KIST
,
R.
и
KISPERT
,
A.
(
9000,
A.
4 ( 9000,
A.
9000 (,
A.
9000 (,
.
Потеря Sox9 в периотической мезенхиме влияет на расширение и дифференцировку мезенхимы, а также на морфогенез эпителия во время развития улитки у мышей
.
Дев. биол.
342
,
51
—
62
.
https://doi.org/10.1016/j.ydbio.2010.03.014
Vincentz
,
J. W.
,
Barnes
,
R. M.
,
Rodgers
,
R
,
Firulli
,
Б.А.0002 Фирулли
,
А. Б.
(
2008
).
Отсутствие Twist1 приводит к аберрантному морфогенезу сердечного нервного гребня
.
Дев. биол.
320
,
131
—
139
.
https://doi.org/10.1016/j.ydbio.2008.04.037
Врличак
,
П.
,
Каллум
,
R.
,
Xu
,
E.
,
Chang
,
A. C. Y.
,
Wederell
,
E. D.
,
Bilenky
,
M.
,
Jones
,
S. J. M.
,
Marra
,
M. A.
,
Karsan
,
A.
и
Без капюшона
,
P. A.
(
2012
).
Транскрипционные мишени Twist1 в развивающемся атриовентрикулярном канале мыши
.
PLoS ONE
7
,
e40815
.
https://doi.org/10.1371/journal.pone.0040815
Вагнер
,
Т.
,
Wirth
,
J.
,
Meyer
,
J.
,
Zabel
,
B.
,
Held
,
M.
,
Zimmer
,
J.
,
Pasantes
,
J.
,
Bricarelli
,
F. D.
,
Keutel
,
J.
,
Hustert
,
E.
и др. (
1994
).
Аутосомная инверсия пола и кампомелическая дисплазия вызываются мутациями в связанном с SRY гене SOX9 и вокруг него
.
Ячейка
79
,
1111
—
1120
.
https://doi.org/10.1016/0092-8674(94)
-8Wheatley
,
S.
,
Wright
,
E.
,
Jeske
,
Y.
,
Mccormack
,
A.
,
Bowles
,
J.
и
Koopman
,
P.
(
1996
).
Этиология синдрома скелетной дисморфологии кампомелической дисплазии: экспрессия Sox9ген во время хондрогенеза у эмбрионов мышей
.
Энн. Н. Я. акад. науч.
785
,
350
—
352
.
https://doi.org/10.1111/j.1749-6632.1996.tb56306.x
Xu
,
N.
,
Lao
,
Y.
,
Zhang
,
Y.
и
Гиллеспи
,
Д. А.
(
2012
).
Akt: палка о двух концах в пролиферации клеток и стабильности генома
.
Дж. Онкол.
2012
,
4
.
https://doi.org/10.1155/2012/
4
Я.
,
Дж.
,
Шилхэм
,
9 0 0 0 1 0 4 Вт.0002 de Boer,
P. A.
,
Moorman
,
A. F. M.
,
Clevers
,
H.
and
Lamers
,
W. H.
(
1998
).
Синдром дефицита Sox4 у мышей является животной моделью общего туловища
.
Обр. Рез.
83
,
986
—
994
.
https://doi.org/10.1161/01.RES.83.10.986
Дополнительная информация
Дополнительная информация- pdf файл
Профилирование одноядерного хроматина вентральной части среднего мозга выявляет факторы транскрипции клеточной идентичности и регуляторные вариации генов, специфичных для типа клеток | Эпигенетика и хроматин
- Исследования
- Открытый доступ
- Опубликовано:
- Yujuan Gui 1 ,
- Kamil Grzyb 2 ,
- Mélanie H. Thomas 2 ,
- Jochen Ohnmacht 1,2 ,
- Pierre Garcia 2 ,
- Manuel Buttini 2 ,
- Александр Скупин 2 ,
- Томас Заутер 1 и
- …
- Лассе Синкконен ORCID: orcid.org/0000-0002-4223-3027 1
Эпигенетика и хроматин том 14 , номер статьи: 43 (2021) Процитировать эту статью
2289 доступов
3 Цитаты
1 Альтметрический
Сведения о показателях
Abstract
Background
Типы клеток в вентральной части среднего мозга участвуют в заболеваниях с различной генетической предрасположенностью, таких как болезнь Паркинсона и шизофрения. Многие генетические варианты влияют на регуляторные области и изменяют экспрессию генов специфичным для типа клеток образом в зависимости от структуры и доступности хроматина.
Результаты
Мы сообщаем о 20 658 профилях доступности одноядерного хроматина вентральной части среднего мозга двух генетически и фенотипически различных линий мышей. Мы различаем десять типов клеток на основе профилей хроматина, а анализ доступных областей, контролирующих гены клеточной идентичности, выделяет ключевые факторы транскрипции, специфичные для типа клеток. Регуляторные вариации, разделяющие линии мышей, проявляются больше на уровне транскриптома, чем на уровне хроматина. Однако данные на уровне клеток показывают изменения, не зафиксированные на уровне тканей. Чтобы узнать масштабы и специфичность клеточного типа цис — действующая вариация в экспрессии генов среднего мозга, мы идентифицируем предполагаемые регуляторные варианты и показываем, что они обогащены дифференциально экспрессируемыми локусами. Наконец, мы обнаружили, что TCF7L2 избирательно опосредует транс -действующие вариации в нейронах среднего мозга.
Выводы
Наш набор данных предоставляет обширный ресурс для изучения регуляции генов в среднем мозге и дает представление о контроле идентичности клеток в среднем мозге и идентифицирует регуляторные вариации, специфичные для типа клеток, которые, возможно, лежат в основе фенотипических и поведенческих различий между линиями мышей.
История вопроса
Вентральный средний мозг, или мезэнцефалон, является одной из наиболее эволюционно законсервированных структур головного мозга млекопитающих [1]. Он участвует в таких задачах, как обработка сенсорной информации и обеспечение моторного и когнитивного контроля через дофаминергические цепи [1]. Он представляет особый интерес в связи с его участием в заболеваниях человека, таких как болезнь Паркинсона и шизофрения, на развитие и прогрессирование которых значительное влияние оказывает индивидуальная генетическая предрасположенность [2,3,4,5].
Как и другие области мозга, средний мозг содержит много различных типов клеток, которые демонстрируют как функциональное, так и молекулярное разнообразие [6,7,8]. Тип клеток можно отличить по профилю экспрессии генов. Транскриптомный анализ на уровне отдельных клеток выявил 20 типов клеток и 58 подтипов в вентральной части среднего мозга [8]. Эти уникальные профили экспрессии генов, определяющие состояние и клеточную идентичность, контролируются эпигенетическими механизмами и достигаются за счет динамического взаимодействия между хроматином и экспрессируемыми транскрипционными факторами (TF). В частности, регуляция с помощью мастер-регуляторов, специфичных для типа клеток, TF, которые открываются и специфически связываются с регуляторными областями генов, приводит к различным профилям экспрессии генов между типами клеток [9].]. Ландшафт хроматина и доступность сайтов связывания TF можно выяснить с помощью эпигеномного анализа, такого как анализ доступного для транспозазы хроматина с последующим секвенированием (ATAC-seq) [10]. До сих пор возможность выделения чистых популяций различных типов клеток мозга ограничивала прогресс в этой области. Однако недавние разработки в анализе одноядерного хроматина в настоящее время сделали возможным массовый параллельный анализ профилей хроматина, специфичных для типа клеток, в их нативном контексте [11,12,13,14].
Типичные геномы человека отличаются друг от друга в среднем на 5 миллионов генетических вариантов [15]. Подавляющее большинство из них расположены в некодирующем геноме, а те, которые связаны со сложными признаками, обогащены в доступных регуляторных областях генов специфичным для типа клеток образом [16]. Генетическая изменчивость в регуляторных регионах может влиять на связывание TF и, таким образом, на экспрессию генов либо в цис , либо в транс , далее именуемые генными регуляторными вариациями [17]. Идентификация генов, регуляторных областей и типов клеток, затронутых регуляторными вариантами, может помочь понять молекулярные механизмы, лежащие в основе рассматриваемого признака. C57BL/6J и A/J представляют собой два генетически различных инбредных штамма мышей, часто используемых в нейробиологии и для изучения сложных генетических признаков. Два штамма разделяются примерно на 6 миллионов вариантов, что сравнимо с генетической изменчивостью между типичными людьми, что делает их интересной модельной системой для понимания влияния регуляторной генетической изменчивости на фенотипическое выражение сложных признаков. Действительно, эти два штамма демонстрируют генетические различия также в чертах, связанных с функцией среднего мозга. Например, A/J более тревожны и менее общительны [18] и имеют более низкую двигательную активность [19].]. Недавно мы показали, что эти два штамма обнаруживают значительные различия в их вентральных транскриптомах среднего мозга [20], но лежащие в их основе регуляторные изменения генов, а также специфичные для типа клеток эпигеномные профили вентрального среднего мозга мышей неизвестны.
Здесь мы выполнили профилирование доступности хроматина вентрального среднего мозга мыши из C57BL/6J и A/J на уровне отдельных ядер (snATAC-seq). Мы идентифицируем> 260 000 отдельных регуляторных областей в 20 658 эпигеномных профилях, которые могут различать десять основных типов клеток в вентральной части среднего мозга, а также определять наборы уникальных генов клеточной идентичности и идентифицировать TF, контролирующие их экспрессию. Сравнение экспрессии генов и доступности хроматина между линиями мышей показывает, что генетически обусловленные различия более выражены на транскриптомном уровне, чем на уровне доступности хроматина. Тем не менее, регуляторные области с альтернативной доступностью хроматина обогащены дифференциально экспрессируемыми генами и могут обнаруживать генную регуляцию, специфичную для типа клеток. Мы находим цис -действующих вариантов, которые должны быть обогащены дифференциально экспрессируемыми генами и точно определять степень регуляторной вариации генов, специфичных для типа клеток. Наконец, мы предполагаем, что каноническая передача сигналов Wnt является медиатором trans -действующих вариаций в нейронах среднего мозга.
Результаты
Профили одноядерного хроматина вентральной части среднего мозга и идентификация основных типов клеток у двух линий мышей
Чтобы разобраться в генной регуляции, специфичной для типа клеток, в среднем мозге и в том, как она влияет на генетические регуляторные вариации, мы провели секвенирование ATAC. на уровне отдельных ядер (snATAC-seq) на выделенном среднем мозге двух генетически различных линий мышей, C57BL/6J и A/J (рис. 1, дополнительный файл 1: рисунок S1). Перфузированные вентральные срезы среднего мозга из двух полушарий одной мыши для обеих линий были использованы для разделения и штрих-кодирования с общим количеством 13 640 и 13 259ядра из C57BL/6J и A/J, соответственно, и после высокопроизводительного секвенирования. После фильтрации мультиплетов и ядер с низким охватом было сохранено приблизительно 290 миллионов прочтений на линию мыши, что соответствует 10 298 (C57BL/6J) и 10 360 (A/J) индивидуальным профилям доступности (рис. 1A). Общий профиль доступности хроматина, агрегированный по отдельным ядрам (массовый snATAC-seq) из C57BL/6J, показал в общей сложности 231 390 пиков. Примечательно, что 99,7% обычных массовых пиков ATAC-seq, полученных из независимого среза среднего мозга C57BL/6J, перекрывались с массовыми пиками snATAC-seq (дополнительный файл 1: рисунок S1). Кроме того, общий профиль snATAC-seq от A/J с 235 157 пиками также сильно коррелировал с профилем C57BL/6J (Pearson R > 0,97). Наконец, чтобы различить доступные области на энхансерах и промоторах, активно участвующих в контроле транскрипции, мы выполнили анализ ChIP-seq на уровне вентральной части среднего мозга у обеих линий мышей на предмет ацетилирования лизина 27 гистона h4 (h4K27ac) [21, 22]. Как массовая ATAC-seq, так и h4K27ac ChIP-seq показали четкую корреляцию с уровнями экспрессии генов среднего мозга (дополнительный файл 1: рисунок S2).
Средний мозг snATAC-seq идентифицирует доступность для конкретного типа клеток. A Кластеризация snATAC-seq из C57BL/6J и A/J с соответствующими пропорциями типов клеток. Основные типы клеток могут быть идентифицированы на основе профилей snATAC-seq, при этом нейроны имеют наибольшую долю в обоих штаммах. Типы клеток в C57BL/6J и A/J имеют сопоставимые пропорции, при этом более половины ядер идентифицируются как нейроны. B Доступность, специфичная для типа клеток, наблюдается в маркерных генах. Геномные треки взяты из snATAC-seq среднего мозга C57BL/6J. Профили экспрессии измерялись как количество транскриптов на 100 000 в кластере. Rpl13a используется в качестве гена домашнего хозяйства для нормализации сигнала snATAC-seq. См. также Дополнительный файл 1: рисунки S1 и S2
Полноразмерное изображение
Используя существующий набор инструментов для геномики отдельных клеток [23, 24], размерность snATAC-seq была рассчитана путем выполнения латентного семантического индексирования (LSI), чтобы обеспечить кластеризацию ячеек с однородной многообразной аппроксимацией и проекцией (UMAP) (рис. 1А). Матрица генной активности snATAC-seq была создана путем подсчета прочтений в теле гена и в промоторной области [2 т.п.о. выше сайта начала транскрипции (TSS)]. Чтобы аннотировать полученные кластеры как отдельные типы клеток, мы воспользовались существующим секвенированием РНК одиночных клеток (scRNA-seq) среднего мозга мыши [8]. Путем идентификации якорных генов, общих для матрицы генной активности snATAC-seq и высоковариабельных признаков в scRNA-seq, мы смогли идентифицировать 10 различных типов клеток среднего мозга с различными профилями доступности хроматина и достаточным количеством клеток в обоих штаммах (рис. 1A). , Дополнительный файл 2: Таблица S1). Типы клеток сгруппированы в шесть основных кластеров, состоящих из глиальных клеток, таких как астроциты (13,5%), микроглия (4,2–5,6%), олигодендроциты (14–19).,5%), и два подтипа полидендроцитов ( Tnr + и Tnr + / Cspg5 + ) (3,2–3,9%), два разных типа эндотелиальных клеток (стеблевые и типичные; 1,1–3,4%), а самые крупные и диффузные кластерные нейроны составляют более половины всех клеток (57,1–60,8%). Хотя данные scRNA-seq позволили различить до 30 подтипов нейронов в среднем мозге посредством комбинаций обилия маркерных генов [8], на уровне доступности хроматина их нельзя было четко различить. Вместо этого можно было четко различить только три класса нейронных клеток: глутаматергические нейроны таламуса (обозначаемые как Slc17a6 + нейронов), дофаминергических нейронов (обозначаемых как Th + нейронов) и более широкой группы нейронов, состоящей в значительной степени, но не исключительно из различных Gad2 + нейронов (ГАМКергических нейронов). называются просто нейронами).
Интересно, что в образцах A/J по сравнению с2116 Tnr + / Cspg5 + полидендроцитов остались практически идентичными (рис. 1А).
Проверка геномных локусов, кодирующих известные маркерные гены, специфичные для клеточного типа, в образцах C57BL/6J, выявила высокую избирательность хроматина в отношении клеточного типа, которая хорошо коррелировала с уровнями экспрессии генов в данных scRNA-seq среднего мозга мыши (рис. 1B). ). В то время как повсеместно экспрессируемый ген Rpl13a имел высокие и постоянные уровни доступности для всех типов клеток, известные маркерные гены для астроцитов ( Aldh2l1 ) и микроглии ( Tmem119 и Selplg ) [25, 26] были экспрессированы и наиболее доступны в соответствующих типах клеток, особенно в их TSS. Точно так же ген, кодирующий переносчик дофамина ( Slc6a3 ), имел самые высокие уровни экспрессии и доступности в нейронах Th + , в то время как в других нейронах сигнал почти не обнаруживался. В то же время соседний ген Clptm1l содержал доступный промотор во всех типах клеток. Наконец, локус, кодирующий два ТФ, необходимых для образования и созревания олигодендроцитов, Olig1 и Olig2 [27, 28] продемонстрировали наибольшую доступность в подтипах полидендроцитов и олигодендроцитов, а также в астроцитах, что опять же соответствует уровням экспрессии генов.
Важно отметить, что профили доступности между C57BL/6J и A/J были в высокой степени сопоставимы также на уровне отдельных типов клеток и могли в равной степени подчеркивать доступность, специфичную для типа клеток, в соответствии с уровнями экспрессии генов, как показано в дополнительном файле 1: рисунок S1 для Aif1 , известный маркерный ген микроглии.
В совокупности наше профилирование snATAC-seq позволило получить более 20 000 профилей хроматина типов клеток среднего мозга мышей сравнимого качества из двух разных линий мышей. Эти данные позволяют различать 10 различных типов клеток среднего мозга на эпигеномном уровне, которые согласуются с известными профилями экспрессии генов.
Идентификация генов клеточной идентичности и связанных с ними регуляторных областей из данных отдельных клеток тип клетки. Чтобы получить эти гены клеточной идентичности, мы использовали существующую scRNA-seq среднего мозга мыши и для каждого гена определили 85-й процентиль его экспрессии во всех типах клеток, указывающий на специфичную для гена «высокую экспрессию» (рис. 2). Чтобы отфильтровать гены, которые избирательно экспрессируются в определенном типе клеток, по крайней мере 60% клеток этого типа клеток имеют экспрессию не ниже 85-го процентиля. Кроме того, для обеспечения уникальности ни одному другому типу клеток не разрешалось иметь один и тот же ген среди наиболее экспрессируемых генов (как определено их 85-м процентилем) более чем в 40% клеток. Благодаря этому подходу мы могли определить от 47 до 412 генов идентичности для каждого типа клеток (рис.
2; дополнительный файл 3: таблица S2). В среднем было определено 170 генов на тип клеток. Чтобы подтвердить соответствие генов биологии рассматриваемого типа клеток, был проведен анализ обогащения GO для биологических процессов. В соответствии с ролью генов в молекулярной и биологической идентичности типов клеток наиболее обогащенные термины GO включали: положительную регуляцию ангиогенеза для клеток эндотелиального стебля; Опосредованный нейтрофилами иммунитет к микроглии; Аксоногенез, транспорт нейротрансмиттеров и регуляция экзоцитоза синаптических пузырьков для различных нейронов; сборка септинового кольца и миелинизация олиго- и полидендроцитов; и негативная регуляция дифференцировки нейронов для астроцитов. Примеры профилей экспрессии генов идентичности из выбранных типов клеток показаны на рис. 2B. Полный список расширенных терминов GO представлен в дополнительном файле 4: Таблица S3. Рис. 2 Области, контролирующие идентичность клеточного типа, можно определить, комбинируя snATAC-seq и scRNA-seq. A Рабочий процесс-схема для определения подписей, специфичных для типа ячейки. Цифровая экспрессия генов получена из DropViz. Для каждого гена рассчитывали 85-й процентиль его экспрессии во всех типах клеток. Чтобы определить ген как ген идентичности клеточного типа, по крайней мере 60% клеток клеточного типа должны иметь экспрессию выше 85-го процентиля, в то же время ни один другой тип клеток не должен иметь такой же ген среди его наиболее экспрессированные гены (выше 85-го процентиля) более чем в 40% клеток. Анализ обогащения генами идентичности клеточного типа обнаружил термины GO, соответствующие характеристикам клеточного типа. Пики идентичности клеточного типа определяются пиками, перекрывающимися с регуляторными областями генов идентичности клеточного типа (базальная область ± 100 т.п.н. до ближайших генов). Впоследствии обнаруживаются обогащенные мотивы в пиках идентичности клеточного типа. B Примеры идентифицированных генов идентичности клеточного типа. Идентифицированные гены идентичности клеточного типа для шести основных типов клеток демонстрируют селективную экспрессию в соответствующих типах клеток при наблюдении за данными scRNA-seq всей популяции клеток среднего мозга
Изображение в натуральную величину
экспрессии генов клеточной идентичности, мы выполнили определение пика для специфичных для типа клеток совокупных сигналов ATAC-seq и связали пики с определенными генами идентичности соответствующих типов клеток, используя GREAT (базальная регуляторная область ± 100 т. п.н. от TSS или до ближайшего ген [29]). В результате было получено 100–1200 доступных областей, которые, вероятно, контролируют экспрессию гена клеточной идентичности в каждом типе клеток (рис. 2; дополнительный файл 5: таблица S4).
Профили доступности хроматина для конкретных типов клеток раскрывают факторы транскрипции, регулирующие клеточную идентичность
Сравнение уровней доступности хроматина по типам клеток подтвердило явное увеличение доступности на полученных пиках клеточной идентичности, связанных с соответствующими генами клеточной идентичности (рис. 3A). ). Наибольшее увеличение сигнала по сравнению с фоном агрегированных клеток среднего мозга всегда обнаруживалось в соответствующих типах клеток, экспрессирующих ассоциированные гены идентичности. В то же время истощение сигнала может быть обнаружено в других типах клеток. Интересно, что уровень доступности также отражал отношения развития типов клеток. Самое сильное истощение сигнала может быть обнаружено в наиболее отдаленном в развитии типе клеток, микроглии [30]. Соответственно, пики идентичности микроглии показали самое сильное истощение во всех других типах клеток. Напротив, пики идентичности нейронов не показали значительного истощения сигнала в родственных Th + нейронов и наоборот. В целом, наш подход может точно идентифицировать регуляторные области генов, специфичных для типа клеток, контролирующих идентичность типа клеток.
Идентификация специфичных для клеточного типа ТФ, контролирующих клеточную идентичность. Тепловая карта , показывающая обогащенный сигнал пиков идентичности типов клеток в восьми типах клеток. Анализ проводили на snATAC-seq среднего мозга C57BL/6J. Фон строится путем слияния считываний выборки (366 278 считываний/тип ячейки) из каждого типа ячеек. Необработанный сигнал нормализуется к фону и библиотеке в соответствии с журналом 9.8579 2 -трансформация. Нормализованный сигнал нанесен на 3 кб выше и ниже пиков. B Анализ обогащения мотивов пиков идентичности клеточного типа. Для каждого мотива показаны логотипы PWM, названия связанных TF и значения p . Мотивы ранжированы в соответствии со значениями p
Изображение с полным размером
Чтобы идентифицировать ТФ, связывающие регуляторные области и контролирующие гены идентичности клеточного типа, мы провели анализ связывающих ТФ мотивов в последовательностях, обогащенных пиками клеточной идентичности (рис. 3B). ). Этот анализ был выполнен для восьми типов клеток с самым высоким охватом секвенирования. Важно отметить, что анализ выявил мотивы для нескольких TF, которые, как ранее было показано, контролируют дифференцировку или идентичность соответствующих типов клеток. К ним относятся SOX9в астроцитах [31], SPI1 в микроглии (известной также как SFPI1 или PU.1 [32, 33]), SOX17 в эндотелиальном стебле [34] и SOX10 и SOX8 в олигодендроцитах [31, 35]. Наиболее богатым мотивом среди типов клеток был общий сайт связывания для CTCF и CTCFL, последовательность, встречающаяся в областях инсулятора, где CTCF опосредует события образования петель хроматина [9], что указывает на образование петель хроматина, селективных по типу клеток, и топологических доменов на участках инсулятора. локусы локусов генов клеточной идентичности.
Кроме того, мотив семейства NFI был обогащен астроцитами и полидендроцитами, что согласуется с его ролью в переходе от нейрогенеза к глиогенезу [36] и необходимостью NFIC для экспрессии маркерных генов астроцитов [37]. Мотивы, обогащенные микроглией, включают мотив MAF, который может связываться с MAFB, TF, который, как недавно было показано, важен для поддержания гомеостаза во взрослой микроглии [38]. Интересно, что мотив семейства RFX был сильно обогащен как в астроцитах, так и в нейронах. Действительно, 9Известно, что 2116 Rfx1 , Rfx3 , Rfx4 и Rfx7 экспрессируются и играют роль в головном мозге [39], причем Rfx4 демонстрирует наибольшую экспрессию в астроцитах, тогда как и Rfx7 Rfx7 многочисленны в различных нейронах [8]. Таким образом, наши данные требуют дальнейшего изучения отдельных ТФ в семействе RFX в клеточной идентичности нейронов среднего мозга и астроцитов.
Вместе с семейством RFX, другой TF с обогащенным мотивом в нейронах, ZNF740, также, как было показано, локализуется в энхансерах генов, активных конкретно в дифференцированных линиях нейронов человека, что еще раз подтверждает актуальность этого предсказания для разных видов [40]. Наконец, мотивы, обогащенные уникальным образом в Th + нейронов включали сайты связывания для TF семейства KLF, TF MEF2 и TF ZBTB7 (которые имеют общий основной мотив с WT1 и ZNF263) (рис. 3B). Из них, в частности, Klf9 , Mef2a , Mef2d и Zbtb7c демонстрируют высокую экспрессию в нейронах Th + [8], при этом Mef2d проявляют более высокую экспрессию, подробные эксперименты по изучению их роли в идентичности дофаминергических нейронов.
Генетически обусловленные изменения доступности хроматина выявляют изменения экспрессии генов, характерные для клеточного типа
Недавно мы показали, что фенотипические различия в среднем мозге между C57BL/6J и A/J (и связанные с ними поведенческие изменения) сопровождаются обширными регуляторными вариациями генов [20 ]. На основании анализа секвенирования РНК на уровне ткани 12 независимых мышей на линию, 1151 ген значительно различается (> в два раза, FDR < 0,05) в вентральной части среднего мозга между двумя линиями (рис. 4A). Тем не менее, 9Механизмы действия 2116 цис и транс , лежащие в основе этих генетически обусловленных изменений, и пораженных типов клеток неизвестны.
Ассоциация дифференциально доступных областей с измененной экспрессией генов между C57BL/6J и A/J. A Дифференциальные пики тесно связаны с дифференциальными генами. Верхние дифференциальные пики (обозначены красным) определяются критерием суммы рангов Уилкоксона с FDR < 0,05 в пределах 1% верхнего значения logFC. Количество прочтений в пиках массива snATAC-seq составляет log 9.8579 10 -преобразованный. Пики с низким числом прочтений (меньше медианы — 1,5 медианы абсолютного отклонения) отфильтровываются. Чтобы связать дифференциальные пики с DEG, пики перекрываются с регуляторной областью DEG (базовая область ± 100 т.п.н. до близлежащих генов). В качестве контроля выбираются случайные пики путем начальной загрузки с 1000 повторений ( p < 0,0099). RPKM из массива РНК-seq C57BL/6J и A/J ( n = 12 на штамм) также является log 10 -трансформированным, а DEG определяется как FDR < 0,05 и log 2 — кратное изменение > 1 (обозначено красным). B Дифференциальные пики, характерные для типов клеток, коррелируют с экспрессией генов в массиве РНК-seq. Дифференциальные пики помечены зеленым цветом. *FDR < 0,05
Полноразмерное изображение
Чтобы оценить вклад изменений уровня хроматина в вариации экспрессии генов, мы сравнили объемные сигналы snATAC-seq от линий мышей и сосредоточились на верхних дифференциальных пиках (FDR < 0,05, верхний 1 % logFC) (рис. 4A). Степень складчатых изменений на уровне доступности хроматина была более скромной, чем на транскриптомном уровне. Тем не менее, 1317 из 263 709названные пики были значительно изменены на объемном уровне. Ассоциируя доступные области с их генами-мишенями, мы могли наблюдать значительное обогащение таких областей дифференциально экспрессируемыми генами. Это указывает на то, что по крайней мере некоторые изменения экспрессии генов могут быть связаны с изменениями на уровне хроматина. Наблюдение за данными на уровне отдельных типов клеток позволило обнаружить дополнительные дифференциально доступные области, специфичные для типов клеток, что указывает на то, что специфичные для типов клеток изменения из редких клеток могут быть замаскированы анализом на уровне тканей. Значительное обогащение дифференциальных пиков дифференциально экспрессируемыми генами сохраняется во всех типах клеток, кроме дофаминергических нейронов, что может быть результатом их низкого охвата секвенированием (рис. 4A).
Для генов, таких как Isoc2b , снижение экспрессии гена в вентральной части среднего мозга A/J сопровождалось снижением доступности промотора во всех типах клеток (фиг. 4B). Чтобы подтвердить, что потеря доступности также сопровождалась снижением транскрипционной активности, мы наблюдали уровни h4K27ac на промоторе. В соответствии со сниженным сигналом ATAC-seq, h4K27ac также был потерян в локусе Isoc2b в A/J.
Для повсеместно экспрессируемых генов, таких как Isoc2b измененная экспрессия гена может быть связана с изменениями уровня хроматина даже при анализе общего уровня. Однако для других генов, таких как Olfr287 , сниженная экспрессия может наблюдаться с помощью секвенирования РНК, хотя на уровне объемного хроматина ни одним из методов (массовым секвенированием ATAC, массовым snATAC-seq и ChIP- послед.). Тем не менее, при наблюдении данных snATAC-seq, специфичных для типа клеток, доступная область может быть обнаружена на промоторе Olfr287 , в частности, в астроцитах. Кроме того, в соответствии со сниженной экспрессией генов хроматин был менее доступен в A/J (рис. 4B).
Таким образом, регуляторные вариации генов в среднем мозге связаны с изменениями доступности уровня хроматина, хотя и не во всех локусах, и с более низкой чувствительностью, чем при транскриптомном анализе. Интересно, что snATAC-seq может выявлять регуляторные изменения, специфичные для типа клеток, которые не фиксируются при анализе объемного уровня.
Предполагаемые
цис -действующие варианты обогащены в регуляторных областях среднего мозга дифференциально экспрессируемыми генами2116 цис — действующая регуляторная вариация, способствующая наблюдаемым различиям в среднем мозге. Для этого мы сосредоточились на идентификации предполагаемых регуляторных вариантов среднего мозга, разделяющих C57BL/6J и A/J. Сначала мы выполнили идентификацию следа TF на нашем профиле доступности хроматина среднего мозга, полученном с помощью массового анализа ATAC-seq. Затем эти следы были перекрыты более чем 6 миллионами вариантов, разделяющих C57BL/6J и A/J, чтобы идентифицировать те, которые могут нарушить связывание TF. Наконец, сайты связывания перекрываются с профилями h4K27ac среднего мозга как C57BL/6J, так и A/J, чтобы захватить сайты связывания, участвующие в транскрипционной активности в любой из линий мышей, что в сумме дает 3909 предполагаемых регуляторных вариантов вентральной части среднего мозга (дополнительный файл 6: таблица S5).
Способность описанного выше подхода уменьшать количество значимых вариантов проиллюстрирована на рис. 5А примерами локусов Ddhd1 , Zfp619 и 4.5S рибосомной РНК (рРНК) . Экспрессия Ddhd1 , гена, кодирующего фосфолипазу, умеренно, но значительно снижена в A/J по сравнению с C57BL/6J и показывает доступный хроматин в его TSS и в расположенном выше энхансерном сайте > 20 кб от TSS. Обе области отмечены сигналами h4K27ac у обоих штаммов. Один след TF может быть идентифицирован как в TSS, так и в дистальном энхансере, что представляет предполагаемые сайты связывания TF, контролирующие Ддхд1 . Из общего числа 603 вариантов в локусе 61 т.п.н. только один совпадает с активным сайтом связывания TF, занимающим средний мозг, что соответствует предполагаемому регуляторному варианту, влияющему на экспрессию Ddhd1 в этой области мозга. Соответственно, пораженный энхансер демонстрирует пониженное обогащение h4K27ac в A/J. Это иллюстрирует, как маловероятно, что большинство генетических вариантов в любом заданном локусе повлияют на экспрессию генов, и как, сосредоточив внимание на тех, которые совместно локализованы в активных регуляторных областях, можно идентифицировать те, которые с наибольшей вероятностью будут действовать как регуляторные варианты.
Предполагаемые регуляторные варианты связаны с дифференциально экспрессируемыми генами и проявляют избирательную доступность в зависимости от типа клеток. A Примеры предполагаемых регуляторных вариантов A/J, обнаруженных в области энхансера выше Ddhd1 , Zfp619 и Rn4.5s . Предполагаемые регуляторные варианты определяются как варианты, нарушающие следы TF, расположенные в активных энхансерах (определяется h4K27ac). B Каждый DEG (5082, FDR < 0,05) связан в среднем с 0,4 предполагаемыми регуляторными вариантами, в то время как не-DEG связаны только с 0,14 вариантами. Случайный: 5000 генов выбираются случайным образом из всех экспрессируемых генов. C Предполагаемые регуляторные варианты имеют разную доступность для разных типов клеток. Более половины вариантов доступны более чем для 6 типов клеток, а 7% — только для 1 типа клеток. D Пример, показывающий, как предполагаемые регуляторные варианты с дифференциальной доступностью влияют на экспрессию генов, специфичных для типа клеток. Варианты, расположенные рядом с TSS Tekt5 , связаны с сигналом TSS в нейронах C57BL/6J, но не A/J, что может привести к усилению регуляции Tekt5 , как показано в массиве РНК-seq. *FDR < 0,05
Изображение полного размера
Если различия в регуляции генов среднего мозга между C57BL/6J и A/J действительно зависят от кумулятивного эффекта цис--действующих вариантов, таких как вариант в локусе Ddhd1 , ожидается, что идентифицированные регуляторные варианты будут обогащены регуляторными областями и сайтами связывания TF в локусах дифференциально экспрессируемых генов по сравнению с другими экспрессируемыми генами. Чтобы проверить это напрямую, мы связали все предполагаемые регуляторные варианты с их вероятными генами-мишенями, как уже показано на рис. 2, и подсчитали количество вариантов, которые в среднем связаны с каждым из 5082 дифференциально экспрессируемых генов (FDR < 0,05) (дополнительный файл 7: Таблица S6) [20]. Мы сделали то же самое для всех экспрессированных генов, которые не изменились между штаммами (FDR > 0,05), и для равного количества случайно выбранных экспрессированных генов в качестве контроля. В то время как незатронутые гены и случайно выбранные гены были связаны в среднем с 0,14 и 0,18 регуляторными вариантами соответственно, это число значительно увеличилось до 0,40 регуляторных вариантов для дифференциально экспрессируемых генов (рис. 5B). Следовательно, генетические варианты, расположенные в регуляторных областях среднего мозга, не обнаруживают случайного распределения, а вместо этого обогащены дифференциально экспрессируемыми генами, предполагая, что они играют важную роль в объяснении наблюдаемых транскриптомных различий.
Затем мы рассмотрели, может ли локализация вариантов в сайтах связывания TF активных энхансеров также быть напрямую связана с активностью энхансера выше изменений экспрессии генов. С этой целью мы использовали THOR [41] для идентификации областей энхансера со значительно измененным сигналом для энхансерной метки h4K27ac между средним мозгом C57BL/6J и A/J. Интересно, что 1126 из 3909 предполагаемых регуляторных вариантов локализованы в энхансерной области с дифференциальным обогащением h4K27ac при строгом пороге (9). 2116 p < 1 × 10 –18 ) (Дополнительный файл 8: Таблица S7). Это указывает на то, что большая часть предполагаемых регуляторных вариантов связана с энхансерами, которые приобретают или теряют активность между линиями мышей. Например, энхансер, содержащий предполагаемые регуляторные варианты в непосредственной близости от локуса, кодирующего 4.5S рРНК , некодирующую РНК, взаимодействующую с рибосомой, демонстрирует сильное усиление энхансерной активности в A/J по сравнению с C57BL/6J (рис. 5A). ). Кроме того, в локусе Zfp619, ген, кодирующий TF цинковых пальцев, как увеличение, так и потерю энхансерной активности можно наблюдать одновременно в двух отдельных энхансерах, связанных с несколькими предполагаемыми регуляторными вариантами. Взятые вместе, нарушение связывания TF вариантами в тысячах областей энхансера, вероятно, изменяет активность энхансера и, таким образом, экспрессию генов среднего мозга у генетически различных линий мышей.
Специфичность к типу клеток
цис--действующих вариантов в среднем мозге Идентификация предполагаемых цис -действующие регуляторные варианты, вносящие вклад в фенотип экспрессии генов среднего мозга между C57BL/6J и A/J, затем мы попытались понять, насколько эти варианты избирательны по типу клеток. Перекрытие предполагаемых регуляторных вариантов с данными о доступности для конкретных типов клеток показало, что большинство сайтов связывания вариантов (57,9%) были доступны и потенциально могли влиять на экспрессию генов по крайней мере в 6 из 10 типов клеток (рис. 5C). . Однако чуть менее 14% вариантов были доступны только в 1 или 2 типах клеток, что указывает на то, что некодирующие вариации могут оказывать селективное влияние на экспрессию генов в зависимости от типа клеток (рис. 5C). Действительно, варианты с доступностью, селективной по типу клеток, только в 1-3 типах клеток значительно чаще встречались в генах с измененной экспрессией, чем в других экспрессируемых генах (данные не показаны).
Ряд регуляторных вариантов был доступен специфически в нейронах и связан с дифференциально экспрессируемыми генами, представляя регуляторную вариацию генов, специфичных для нейронов. К ним относятся, например, Tekt5 , ген, экспрессируемый в возбуждающих нейронах и активируемый в C57BL/6J (рис. 5D). Несмотря на наблюдаемую дифференциальную экспрессию, промотор Tekt5 оказался недоступным в большинстве типов клеток, за исключением нейронов C57BL/6J, именно при перекрытии TSS с предполагаемым регуляторным вариантом в пределах сайта связывания TF.
В совокупности, хотя большинство цис -действующих вариантов воздействуют на широкий спектр типов клеток, большая их часть может иметь эффекты, специфичные для типа клеток, которые невозможно выявить без анализа отдельных клеток.
TCF7L2 как медиатор
транс--действующих вариаций в нейронах среднего мозга След ТФ расположен в активном энхансере (рис. 5А). Тем не менее, большая часть дифференциальной экспрессии генов оставалась необъясненной. Это может быть из-за цис--действующие варианты, которые мы пропустили, но также из-за транс--действующих вариантов, которые могут влиять на ряд генов-мишеней, изменяя активность ТФ, а не мотив его связывания. Изменение активности TF может быть связано с изменением уровня его экспрессии, но также может быть связано с чередованием других механизмов, контролирующих активность TF, таких как посттрансляционные модификации, белок-белковые взаимодействия или локализация TF. Генетические различия в недофаминергических нейронах (например, Gad2 + нейронов), которые составляют большую часть нашей популяции нейронов (дополнительный файл 2: таблица S1), предположительно вносят вклад в специфические поведенческие различия, включая тревожность, вознаграждение и мотивационные черты, такие как потребление этанола. 42,43,44,45]. Чтобы идентифицировать медиаторы транс--действующих вариаций между C57BL/6J и A/J в нейронах, мы провели анализ обогащения мотивов на 715 участках нейронов, демонстрирующих разную доступность хроматина у мышей (рис. 4A). Это показало, что мотив связывания семейства TCF/LEF, расположенных ниже TF канонического сигнального пути Wnt [46], входит в число наиболее обогащенных последовательностей, обнаруженных в дифференциально доступных областях (рис. 6A; 9).2116 p = 1,66e−14). Обогащение было специфичным для Gad2 + нейронов и не могло быть обнаружено ни в каких других типах клеток (дополнительный файл 1: рисунок S3) или в анализе обогащения мотивов для генов клеточной идентичности (рис. 3). LEF1, TCF7L1 и TCF7L2 связываются с одной и той же последовательностью ДНК, но часто имеют противоположные или специфичные для типа клеток функции [47]. Проверка доступности хроматина в разных типах клеток для дифференциальных сайтов связывания, несущих мотив TCF/LEF, выявила повышенный сигнал именно в нейронах (рис. 6B). Чтобы определить, какой фактор экспрессируется в нейронах среднего мозга и может опосредовать наблюдаемое обогащение и измененную доступность, мы визуализировали их экспрессию с использованием данных scRNA-seq. 9Экспрессия 2116 Lef1 была ограничена эндотелиальными клетками (рис. 6C), а Tcf7l1 показала только низкую экспрессию или отсутствие экспрессии во всех типах клеток (рис. 6D). Однако Tcf7l2 имел высокую экспрессию в нейронах и полидендроцитах Gad2 + и Slc17a6 + , демонстрируя явное перекрытие с клетками, обогащенными соответствующим мотивом связывания (рис. 6E). Соответственно, анализ предполагаемых вышестоящих регуляторов, объясняющих транскриптомные изменения из секвенирования РНК мышей с использованием анализа пути изобретательности (IPA), предсказал, что TCF7L2 (также известный как TCF4) и CTNNB1 (бета-катенин, партнер по связыванию TCF7L2) будут среди ведущие регуляторы на основе прогнозируемой оценки активации (Дополнительный файл 9: Таблица S8). Таким образом, транскрипционная активность TCF7L2, вероятно, изменяется между мышами C57BL/6J и A/J в нейронах среднего мозга.
Семейство TCF/LEF как медиатор транс -действующих вариаций в нейронах. A Анализ обогащения мотивов для областей дифференциальной доступности между C57BL/6J и A/J в нейронах. B , C Области дифференциальной доступности с мотивом семейства TCF/LEF обнаруживают повышенную доступность в нейронах по сравнению с другими типами клеток. D – E Lef1 высоко экспрессируется в эндотелиальном стебле , Tcf7l1 демонстрирует низкую экспрессию во всех типах клеток, в то время как Tcf7l2 в изобилии присутствует в нейронах и полидендроцитах. См. также Дополнительный файл 1: Рисунок S3
Полноразмерное изображение
В совокупности анализ snATAC-seq тканей из генетически различных штаммов может помочь в выяснении специфического для клеточного типа влияния транс--действующих вариантов и позволяет предположить нейрон- специфические различия в каноническом сигнальном пути Wnt между двумя обычно используемыми линиями инбредных мышей.
Обсуждение
Здесь мы исследовали доступность хроматина типов клеток вентрального среднего мозга мыши в двух различных генетических фонах и предоставили большой ресурс из 20 658 профилей одноядерного хроматина на 10 различных типах клеток. Этот набор данных принесет пользу будущим исследованиям роли этих типов клеток в различных процессах, связанных с вентральной частью среднего мозга, таких как управление движением, познание и механизмы вознаграждения. Лучшее понимание типов клеток среднего мозга также может помочь в исследованиях таких заболеваний, как болезнь Паркинсона и шизофрения. В частности, лучшее понимание генетической изменчивости в регуляции генов и того, как она влияет на определенные типы клеток, проложит путь к более точному прогнозированию генетической предрасположенности и механизмов пораженных заболеваний.
Наши данные о профилях доступности генов и составе клеток среднего мозга согласуются с существующими знаниями по транскриптомике одиночных клеток [8]. Однако подтипы нейронов труднее различить на уровне хроматина, чем это было достигнуто с помощью транскриптомного анализа. Подтипы нейронов сгруппированы в основном вместе, часто с неопределенными границами между подтипами (рис. 1А). Этот результат ожидаем. Хотя общеизвестно, что доступность хроматина показывает положительную корреляцию с экспрессией генов [48, 49], и это также верно для наших данных (дополнительный файл 1: рисунок S2), доступность энхансера не обязательно отражает активность регуляции генов [50]. Действительно, профили доступности хроматина типов клеток, выполняющих сходные функции, могут быть очень сходными, несмотря на то, что демонстрируют разные паттерны экспрессии и контролируются разными мастер-TF [16, 51]. Более того, доступность может означать праймирование локуса для экспрессии без начала транскрипции [52, 53].
Тем не менее, мы смогли надежно различить 10 из 20 известных типов клеток среднего мозга на уровне хроматина, при этом восемь типов клеток содержали достаточное количество клеток для детального анализа (рис. 3). Используя информацию о регуляторных областях генов, избирательно связанных с генами, лежащими в основе клеточной идентичности, мы смогли предсказать ключевые регуляторы каждого типа клеток с помощью анализа обогащения мотивов. К ним относятся многие факторы, которые ранее считались необходимыми для дифференцировки или поддержания соответствующего клеточного состояния [31, 32, 33, 35, 54]. Помимо прочего, результаты показывают специфичную для типа клеток роль сайтов связывания CTCF в клеточной идентичности и предсказывают специфическую роль TF семейства MEF2, KLF и ZBTB7 в дофаминергических нейронах. Более того, результаты дают дополнительную поддержку для более детального анализа семейства TFs RFX в регуляции баланса нейро- и глиогенеза в среднем мозге, что согласуется с ранее идентифицированным совместным присутствием факторов RFX в SOX2-связанных энхансерах в развитии нервной системы. 55].
Известно, что генетическая изменчивость вызывает изменения экспрессии генов, которые могут вызывать фенотипические различия [56,57,58,59]. Недавно мы показали, что более тысячи генов по-разному экспрессируются в среднем мозге мышей из-за генетической изменчивости между C57BL/6J и A/J [20]. Здесь мы использовали наши профили доступности хроматина отдельных клеток для исследования механизмов, специфичности клеточного типа и степени, в которой это изменение отражается на уровне хроматина. Интересно, что транскриптомные различия не показали линейной зависимости от доступности хроматина среднего мозга (рис. 4А). Степень изменения доступности несопоставима с изменением уровня мРНК ни на уровне объемных агрегатов, ни в отдельных типах клеток. Скорее всего, это отражает наблюдение, что связывание и активность TF могут изменять экспрессию генов без изменения доступности, например, если локус уже открыт [60]. Тем не менее, когда изменения доступности действительно происходили, это часто было связано с изменением экспрессии локального гена. Важно отметить, что некоторые изменения доступности хроматина, связанные с дифференциальной экспрессией генов, вообще не могли быть обнаружены при анализе ATAC-seq или ChIP-seq на уровне ткани, несмотря на большую глубину секвенирования (рис. 4B). Таким образом, улучшенное разрешение, обеспечиваемое анализом отдельных клеток, позволяет лучше понять регуляторные взаимодействия, которые могут быть упущены при анализе на уровне ткани.
Cis -действующие варианты, которые влияют на экспрессию генов, были предложены для объяснения значительной части отсутствующей наследуемости [61,62,63]. В настоящее время экспериментально подтверждено растущее число таких вариантов, связанных с человеческими чертами и заболеваниями [17]. Объединив генетическую информацию с анализом ATAC-seq и ChIP-seq, мы обнаружили, что 3909 из > 6 миллионов вариантов, разделяющих C57BL/6J и A/J, локализуются в сайте связывания TF в активном и доступном энхансере в вентральной части среднего мозга (рис. 5). Это число согласуется с предыдущими оценками, согласно которым один вариант из тысячи мышей вызывает цис — регуляторные эффекты [64]. Важно отметить, что мы обнаружили, что предполагаемые цис--действующие варианты обогащены дифференциально экспрессируемыми генами, что указывает на то, что они действительно вносят вклад в фенотип экспрессии генов.
Большая часть предполагаемых цис -действующих вариантов показала избирательную доступность по типу клеток (рис. 5B). Этот вывод согласуется с предыдущей работой по влиянию цис -действующих вариантов на связанные с заболеванием регуляторные изменения в основных клеточных популяциях головного мозга [65]. Объединение информации о влиянии вариантов на регуляцию генов, специфичных для типа клеток, с полногеномными исследованиями ассоциаций и количественным картированием локусов определенных признаков или заболеваний может дать представление о том, как варианты трансформируются в фенотипы. C57BL/6J и A/J отличаются друг от друга многочисленными фенотипами [66]. К ним относятся поведение, обусловленное страхом и вознаграждением, такое как потребление этанола, которое связано с передачей сигналов дофамина в среднем мозге [43,44,45]. Объединяя наши данные о селективности клеток цис -действующие варианты с полногеномными исследованиями ассоциаций таких фенотипов могут выявить новые связи между генами и лежащими в их основе механизмами.
Помимо цис--действующей вариации, дифференциальная экспрессия генов и измененная доступность хроматина также могут быть опосредованы посредством транс--действующей вариации. Транс--действующие варианты могут иметь множество различных механизмов действия, и было высказано предположение, что большая часть генетически обусловленных вариаций экспрессии генов связана с транс -действующая вариация, часто затрагивающая сотни или тысячи генов клеточно-специфическим образом [62, 67, 68]. Дофаминергические нейроны в вентральной области покрышки среднего мозга вносят вклад в фенотипы, такие как поведение вознаграждения и страх [42]. Более того, ассоциированные ГАМКергические и глутаматергические нейроны в среднем мозге также могут контролировать такое поведение либо вместе с передачей сигналов дофамина, либо независимо от нее. Имея это в виду, мы спросили, можно ли обнаружить специфические признаки транс--действующих регуляторных вариаций в нейронах между нашими штаммами, которые демонстрируют дифференциальное поведение, на основе анализа обогащения мотивов в областях дифференциальной доступности (рис. 6). Мы обнаружили обогащение мотива связывания TCF/LEF, соответствующего элементу ответа на сигнализацию Wnt, распознаваемому их ДНК-связывающим доменом группы высокой подвижности (HMG) [46]. Это предполагает, что передача сигналов Wnt изменяется между C57BL/6J и A/J в нейронах. На основании данных scRNA-seq, TCF7L2, но не LEF1 или TCF7L1, обильно экспрессируется в среднем мозге Gad2 + и Slc17a6 + нейронов, что указывает на то, что изменения доступности хроматина и экспрессии генов связаны с изменениями в передаче сигналов Wnt и, вероятно, опосредованы TCF7L2 (рис. 6). Действительно, анализ путей дифференциально экспрессируемых генов выявил обогащение известных генов-мишеней Wnt среди DEG (дополнительный файл 9: таблица S8).
Участие TCF7L2 в качестве предполагаемого эффектора измененной передачи сигналов в нейронах особенно интригует, поскольку 9Локус 2116 TCF7L2 был генетически связан с психическими расстройствами, такими как шизофрения и биполярное расстройство у людей [69, 70]. Более того, у трансгенных мышей выявлена дозозависимая роль Tcf7l2 в формировании страха и тревоги, признаков, по которым A/J, как известно, значительно отличается от C57BL/6J [43, 44]. Интересно, что влияние дозы Tcf7l2 на ряд фенотипов, включая поведенческие фенотипы, как известно, зависит от генетического фона мыши [71]. В то время как некоторые фенотипы, вызванные снижением 9Экспрессия 2116 Tcf7l2 может наблюдаться на фоне A/J, но не на фоне C57BL/6J, противоположное верно для некоторых других фенотипов. Основываясь на наших данных, одним из возможных объяснений этого может быть роль Tcf7l2 в качестве посредника транс -действующей вариации и, таким образом, дифференциальная базовая активность пути Wnt между линиями мышей. Понимание того, как активность передачи сигналов Wnt изменяется между линиями мышей, требует дальнейшего анализа. Однако интересно отметить, что выражение Wnt2b , восходящий лиганд пути Wnt, также генетически связанный с биполярным расстройством [69], значительно снижен в вентральной части среднего мозга A/J по сравнению с C57BL/6J [20].
Выводы
Взятые вместе, наш анализ одноядерного хроматина обеспечивает новое понимание транскрипционного контроля вентральных типов клеток среднего мозга и богатый ресурс для дальнейшего анализа клеточной идентичности и генных регуляторных вариаций в этой области мозга, связанной с заболеванием.
Методы
Животные
Все экспериментальные процедуры в этом исследовании проводились в соответствии с Директивой Совета Европейского сообщества 2010/63/ЕС в соответствии с требованиями 3 Rs в отношении благополучия животных. В этом исследовании мы использовали две линии мышей, C57BL/6J и A/J, приобретенные, соответственно, у Charles River and Jackson Laboratory. Когорты исследования были выведены в Центре животных Люксембургского университета (Эш-сюр-Альзетт, Люксембург), и протокол был одобрен Комитетом по этике экспериментов на животных (AEEC) в соответствии с национальными рекомендациями закона о защите животных в Люксембург ( 9Великокняжеский Постановление № 2116 № от 11 января 2013 г. ). Все мыши содержались при 12-часовом режиме свет-темнота и имели свободный доступ к пище и воде.
Для каждой мыши после анестезии смесью кетамина и медетомидина (150 и 1 мг/кг соответственно) выполняли внутрисердечную перфузию с помощью PBS. Мозг извлекали, и оба среднего мозга каждой мыши препарировали, немедленно замораживали, хранили при температуре — 80 °C и использовали для разделения отдельных клеток, как описано ниже.
Выделение ядер
Для нерасфасованного ATAC-seq замороженный средний мозг измельчали в измельчителе тканей Dounce (Sigma, D8939-1SET) с пестиком A ~ 10 раз после пестика B ~ 20 раз в лизирующем буфере [5 мМ CaCl 2 (Merck, A546282 ), 3 мМ Mg(Ac) 2 (Roth, P026.1), 10 мМ Трис, pH 7,8, 0,17 мМ β-меркаптоэтанол (Gibco, 21985-023), 160 мМ сахарозы (Sigma, S0389), 0,05 мМ ЭДТА , 0,05% NP40 (Sigma, I3021)]. Лизат наслаивали на 3 мл сахарозной подушки [1,8 М сахарозы, 3 мМ Mg(Ac)2, 10 мМ Трис, pH 8,0, 0,167 мМ β-меркаптоэтанола] после ультрацентрифугирования 30 000 г в течение 1 ч (ротор: Beckman Coulter, MLS-50) при 4 °C. После центрифугирования супернатант удаляли, а осадок ядер суспендировали в буфере для ресуспендирования (с 0,1% Tween-20, 0,01% дигитонина и 0,1% NP40).
Для snATAC-seq выделение ядер проводили в соответствии с протоколом 10× Genomics CG000212 (Rev A) с небольшими изменениями. Для обоих штаммов срезы среднего мозга из двух полушарий одной и той же мыши обрабатывали отдельно и объединяли на уровне анализа. Вкратце, замороженные срезы среднего мозга измельчали в измельчителе тканей Dounce (Sigma, D89).39-1SET) с пестиком A ~ 10 раз после пестика B ~ 20 раз в 500 мкл охлажденного буфера для лизиса [10 мМ Трис-HCl (pH 7,4), 10 мМ NaCl, 3 мМ MgCl 2 , 0,1% Tween-20 , 0,1% NP40, 0,01% дигитонин, 1% БСА]. Гомогенизированный лизат инкубировали на льду в течение 5 мин, после чего перемешивали пипеткой 10 раз и снова инкубировали на льду в течение 10 мин. К лизированным клеткам добавляли охлажденный промывочный буфер (500 мкл) [10 мМ Tris-HCl (pH 7,4), 10 мМ NaCl, 3 мМ MgCl 2 , 1% BSA, 0,1% Tween-20] и пипетировали смесь 5 раз. . Лизат наслаивали на 3 мл сахарозной подушки [1,8 М сахарозы, 3 мМ Mg(Ac)2, 10 мМ Трис, pH 8,0, 0,167 мМ β-меркаптоэтанола] после ультрацентрифугирования 30 000×9.2116 г в течение 1 ч (ротор: Beckman Coulter, MLS-50) при 4 °C. После центрифугирования надосадочную жидкость удаляли, а осадок ядер суспендировали в буфере ядер (10× PN: 2000153), входящем в комплект 10× Genomics Chromium Single Cell ATAC Reagent Kit. Суспензию ядер пропускали через 40-мкм клеточный фильтр Flowmi (Sigma, BAh236800040-50EA).
Bulk ATAC-seq
Маркировку образца среднего мозга мыши C57BL/6J проводили на основании дополнительного протокола 1 OMNI-ATAC [72] с небольшими изменениями. Вкратце, 25 000 ядер ресуспендировали в 50 мкл буфера для ресуспендирования (с 0,1% Tween-20, 0,01% дигитонина и 0,1% NP40) и лизировали в течение 3 минут на льду. После промывки в 1 мл буфера для ресуспендирования (1% Tween-20) образцы центрифугировали в течение 10 мин при 500× г при 4 °C и осторожно удаляли надосадочную жидкость. Осадки ресуспендировали в 25 мкл смеси для мечения (буфер для ДНК-метки от Illumina, № 15027866), содержащей 2,5 мкл фермента ДНК-метки (Illumina № 15027865), и инкубировали в течение 45 минут при 37 °C и 1000 об/мин в Eppendorf ThermoMixer. Меченый хроматин выделяли с использованием набора Zymo Research DNA Clean & Concentrator (ZymoResearch ZY-D4013) и элюировали в 21 мкл элюирующего буфера. Предварительную амплификацию библиотеки проводили в течение 5 циклов с использованием праймеров Ad1 и Ad2.16 [10]. Было проведено пять дополнительных циклов амплификации библиотеки, как определено с помощью количественной ПЦР [72]. Очистку библиотеки проводили с использованием набора Zymo Research DNA Clean & Concentrator с последующим выбором размера гранул AMPure XP (Beckman Coulter #A63880). Первая инкубация шариков с использованием 0,55-кратного объема шариков удаляет большие фрагменты. После разделения шариков на магнитной подставке супернатант переносили в свежую пробирку и инкубировали в течение 5 мин в 1,5-кратном объеме шариков. После промывания 80% этанолом шарики ресуспендировали в 20 мкл элюирующего буфера. После разделения на магнитном штативе элюат переносили в свежую пробирку. Качество библиотеки оценивали с помощью высокочувствительного биоанализатора Agilent DNA (Agilent № 5067-4626).
Массовый анализ данных ATAC-seq
Секвенирование библиотек ATAC-seq было выполнено на платформе секвенирования в Люксембургском центре системной биомедицины (LCSB) Люксембургского университета. Секвенирование парных концов нецепочечной библиотеки проводили с использованием набора Illumina NextSeq 500/550 75 циклов High Output Kit. Необработанные файлы FASTQ и файлы BAM обрабатывали, как описано в разделе «Методы» для ChIP-seq. После обработки файлов BAM пики были вызваны Генрихом (https://github.com/jsh58/Genrich) с параметрами «-r -m 30 -j» для удаления дубликатов ПЦР и включения только чтений с качеством отображения на уровне не менее 30. Футпринты вызывались HINT-ATAC [73] с параметрами по умолчанию. Необработанные файлы FASTQ были депонированы в ArrayExpress под регистрационным номером E-MTAB-8333.
Подготовка библиотеки одноядерных ATAC-seq и секвенирование
Одноядерный ATAC-seq выполняли в соответствии с Руководством пользователя наборов реагентов Chrominum Single Cell ATAC Reagent Kit (CG000168 RevB) с использованием набора Chromium Single Cell E Chip Kit (10×, 100086). , Chromium Single Cell ATAC Library & Gel Bead Kit (10×, 1000111), Chromium Single Cell ATAC Library Kit (10×, 1000085), Chromium Single Cell ATAC Library Kit (10× 100087), Chromium i7 Multiplex Kit N Set ( 1000084), Dynabeads MyOne Silane (2000048). Вкратце, суспензию ядер загружали с целевой скоростью извлечения 10 000 ядер на образец. Качество библиотек snATAC-seq оценивали с использованием чипа высокочувствительного биоанализатора ДНК Agilent (Agilent № 5067-4626) и далее секвенировали на наборе High Output Kit на 150 циклов с использованием Illumina NextSeq™ 500 с заданной глубиной секвенирования 25 000 пар прочтений на ядро. Необработанные файлы FASTQ были депонированы в ArrayExpress под номером доступа E-MTAB-9. 225.
Конвейер анализа ATAC-seq для отдельных ядер
Cell Ranger
Выравнивание и фильтрацию проводили в соответствии с 10-кратными конвейерами. Вкратце, файлы fastq были сгенерированы из файлов базовых вызовов секвенсора Illumina, которые позже использовались в качестве входных данных для выравнивания (MAPQ > 30), фильтрации мультиплетов штрих-кода и создания счетчиков доступности для каждой ячейки в одной библиотеке. Технические повторы для каждой линии мышей были объединены для создания единой матрицы пикового штрих-кода. Каждый уникальный фрагмент связан со штрих-кодом одной ячейки.
Кластеризация
Нисходящий анализ snATAC-seq был выполнен компанией Signac (версия: 0.2.4) в R. Матрица генной активности была рассчитана с считываниями в теле гена и 2 т.п.н. выше TSS в качестве показателя экспрессии гена. Ядра с числом менее 5000 отфильтровывали. Размерность рассчитывалась с помощью латентного семантического индекса (LSI) на пиках с не менее чем 100 считываниями во всех ячейках, которые использовались в качестве входных данных для создания графиков UMAP.
Аннотация кластера
При аннотации кластеров snATAC-seq использовались существующие данные scRNA-seq по среднему мозгу взрослых трехмесячных мышей C57BL/6N. Якоря были обнаружены между матрицей генной активности snATAC-seq и 5000 лучших вариабельных признаков scRNA-seq. Клеточные метки были перенесены из scRNA-seq в snATAC-seq с нормализацией по весам якорей, рассчитанным на основе уменьшения размеров LSI, в результате чего было аннотировано 10 347 ядер C57BL/6J и 10 368 ядер A/J.
Конвейер анализа данных scRNA-seq
Обработка данных
Цифровая экспрессия генов (DGE) и аннотация клеток для среднего мозга 3-месячных мышей C57BL/6N были загружены с DropViz. Анализ данных был выполнен Seurat (версия: 3.1.4) в R. В последующем анализе использовались только ячейки с числом признаков от 400 до 7000, которые были одиночными или тщательно отобранными, в результате чего было получено 19 967 ячеек. DGE был преобразован в натуральный логарифм и нормализован по количеству митохондриальных прочтений. Уменьшение размеров было выполнено с помощью UMAP. Кластеры были аннотированы существующей аннотацией от DropViz.
Отбор генов идентичности типа клеток
Из каждого клеточного кластера случайным образом отобрали 100 клеток. DGE из каждого типа клеток конструировали по соответствующим штрих-кодам. Для выбранных клеток рассчитывали 85-й процентиль экспрессии для каждого гена. Критерии генов идентичности клеточного типа были определены следующим образом: для конкретного гена 60% клеток клеточного типа имеют экспрессию, превышающую специфичный для гена 85-й процентиль; в то время как не более 40% клеток всех других типов клеток имеют экспрессию выше 85-го процентиля. Этот процесс повторялся 100 раз. Гены, которые появлялись более 30 раз из 100, были определены как гены идентичности клеточного типа.
Определение дифференциальных пиков
Дифференциальные пики определялись для каждого типа клеток с использованием критерия суммы рангов Уилкоксона (реализовано в пакете R «presto» 1. 0.0). Матрица подсчета двоичных пиков из scATAC-seq была нормализована по количеству пиков, представленных в каждой ячейке. Затем был проведен дифференциальный анализ пиков с presto на нормализованной матрице подсчета пиков. Пики с FDR < 0,05 и логарифмической кратностью изменения (logFC) в верхнем 1% определяются как дифференциальные пики.
Анализ обогащения мотивов
Создание файлов bam для конкретных типов ячеек
Файлы bam для конкретных типов ячеек были созданы программой samtools. Штрих-коды из каждой реплики штамма в bamfile были перемаркированы, чтобы избежать коллапса штрих-кода. После перемаркировки бам-файлы из повторов образца были объединены. Бам-файлы для каждого типа клеток вычитались на основе соответствующих штрих-кодов.
Пиковый вызов
Пиковый вызов был выполнен MACS2 (2.1.2) [74] с настраиваемой отсечкой по p значений в соответствии с анализом отсечки с параметрами «macs2 callpeak -cutoff-analysis». Идеальная отсечка была выбрана на основании выбранных p значение не приведет к экспоненциальному увеличению числа пиков.
Анализ обогащения мо
Иммунопреципитация хроматина (ChIP)
ChIP была проведена на рассеченной мгновенно замороженной ткани вентральной части среднего мозга двух независимых мышей каждой линии, как описано ранее [20]. В каждой реакции было 10–14 мкг хроматина, и в качестве входной ДНК использовалась 10% аликвота. Иммунопреципитацию проводили в течение ночи с 5 мкл антитела h4K27ac (Abcam, ab4729), при 4 °C с вращением.
Анализ данных ChIP-seq
Секвенирование образцов хроматина было выполнено на платформе секвенирования в LCSB Университета Люксембурга. Одностороннее нецепочечное секвенирование с длиной считывания 75 п.н. было выполнено на машине Illumina NextSeq 500. FastQC (v0.11.5) использовался для оценки качества необработанных прочтений [76]. Генерация файлов BAM, включая этапы удаления адаптера, сопоставления и маркировки дубликатов, выполнялась с помощью конвейера PALEOMIX (v1.2.12) [77], после чего выполнялось сопоставление с помощью BWA (v. 0.7.16a) [78]. Алгоритм Backtrack применил смещение качества оценки Phred до 33. Дубликаты чтения были отмечены, и эталонный геном мыши, GRCm38.p5 (mm10, патч 5), был загружен из GENCODE (https://www.gencodegenes.org/). Наконец, проверка файлов Bam была выполнена с помощью Picard (v2.10.9).) [79]. Необработанные файлы FASTQ были депонированы в ArrayExpress под номером доступа E-MTAB-8333.
Пики h4K27ac ChIP-seq, энхансеры и суперэнхансеры были вызваны HOMER [75] с параметрами по умолчанию. Нормализация сигнала при попарном сравнении выполнялась с помощью THOR (v0.10.2) [41] с нормализацией TMM и скорректированным значением отсечки p 0,01.
Доступность данных и материалов
Необработанные файлы FASTQ были депонированы в ArrayExpress под номером доступа E-MTAB-8333 для анализа объемных уровней и E-MTAB-9225 для данных одной ячейки. Треки snATAC-seq доступны в браузере генома UCSC (http://genome-euro.ucsc.edu/cgi-bin/hgTracks?hubUrl=https://biostat2.uni.lu/ygui/hub. txt&genome=mm10). ). Доступ к скриптам для анализа данных можно получить здесь: https://github.com/sysbiolux/Gui-et-al.
Ссылки
Vogt Weisenhorn DM, Giesert F, Wurst W. Разнообразие имеет значение — гетерогенность дофаминергических нейронов в вентральном мезэнцефалоне и его связь с болезнью Паркинсона. Дж. Нейрохим. 2016;139(Приложение 1): 8–26.
КАС пабмед ПабМед Центральный Статья Google ученый
Кляйн С., Вестенбергер А. Генетика болезни Паркинсона. Колд Спринг Харб Перспект Мед. 2012;2(1):a008888.
ПабМед ПабМед Центральный Статья Google ученый
Li Z, Chen J, Yu H, He L, Xu Y, Zhang D и др. Полногеномный ассоциативный анализ выявил 30 новых локусов предрасположенности к шизофрении. Нат Жене. 2017;49(11): 1576–83.
КАС пабмед Статья Google ученый
Наллс М.А., Блаувендраат С., Валлерга С.Л., Хейлброн К., Бандрес-Сига С., Чанг Д. и др. Выявление новых локусов риска, понимание причин и наследуемый риск болезни Паркинсона: метаанализ ассоциативных исследований по всему геному. Ланцет Нейрол. 2019;18(12):1091–102.
КАС пабмед ПабМед Центральный Статья Google ученый
Уильямс Х.Дж., Оуэн М.Дж., О’Донован М.С. Новые результаты исследований генетической ассоциации шизофрении. Джей Хам Жене. 2009;54(1):9–14.
КАС пабмед Статья Google ученый
Gantz SC, Ford CP, Morikawa H, Williams JT. Развивающееся понимание дофаминовых нейронов в черной субстанции и вентральной области покрышки. Annu Rev Physiol. 2018;80(1):219–41.
КАС пабмед Статья Google ученый
Сондерс А., Макоско Э.З., Высокер А., Голдман М., Кринен Ф.М., де Ривера Х. и др. Молекулярное разнообразие и специализации клеток мозга взрослых мышей. Клетка. 2018;174(4):1015-1030.e16.
КАС пабмед ПабМед Центральный Статья Google ученый
Atlasi Y, Stunnenberg HG. Взаимодействие эпигенетических меток во время дифференцировки и развития стволовых клеток. Нат Рев Жене. 2017;18(11):643–58.
КАС пабмед Статья Google ученый
Buenrostro J, Wu B, Chang H, Greenleaf W. ATAC-seq: метод анализа доступности хроматина для всего генома.
Curr Protoc Mol Biol Ed Frederick M Ausubel Al. 2015;109:21.29.1-21.29.9.Google ученый
Кусанович Д.А., Хилл А.Дж., Агамирзае Д., Даза Р.М., Плинер Х.А., Берлетч Д.Б., и др. Одноклеточный атлас доступности хроматина млекопитающих in vivo. Клетка. 2018;174(5):1309-1324.e18.
КАС пабмед ПабМед Центральный Статья Google ученый
Lake BB, Chen S, Sos BC, Fan J, Kaeser GE, Yung YC и др. Интегративный одноклеточный анализ транскрипционных и эпигенетических состояний во взрослом мозге человека. Нац биотехнолог. 2018;36(1):70–80.
КАС пабмед Статья Google ученый
Preissl S, Fang R, Huang H, Zhao Y, Raviram R, Gorkin DU, et al. Одноядерный анализ доступного хроматина в развивающемся переднем мозге мышей выявляет специфичную для типа клеток регуляцию транскрипции.
Нат Нейроски. 2018;21(3):432–9.КАС пабмед ПабМед Центральный Статья Google ученый
Sinnamon JR, Torkenczy KA, Linhoff MW, Vitak SA, Mulqueen RM, Pliner HA, et al. Доступный хроматиновый ландшафт мышиного гиппокампа при разрешении одной клетки. Геном Res. 2019;29(5):857–69.
КАС пабмед ПабМед Центральный Статья Google ученый
Auton A, Abecasis GR, Altshuler DM, Durbin RM, Abecasis GR, Bentley DR, et al. Глобальный справочник по генетической изменчивости человека. Природа. 2015;526(7571):68–74.
ПабМед Статья КАС Google ученый
Maurano MT, Humbert R, Rynes E, Thurman RE, Haugen E, Wang H, et al. Систематическая локализация распространенных при заболеваниях вариаций в регуляторной ДНК. Наука. 2012;337(6099): 1190–5.
КАС пабмед ПабМед Центральный Статья Google ученый
Депланке Б., Альперн Д., Гарде В. Генетика вариаций связывания ДНК фактора транскрипции. Клетка. 2016;166(3):538–54.
КАС пабмед Статья Google ученый
Мой С.С., Надлер Дж.Дж., Янг Н.Б., Перес А., Холлоуэй Л.П., Барбаро Р.П. и др. Поведенческие задачи мышей, имеющие отношение к аутизму: фенотипы 10 инбредных штаммов. Поведение мозга Res. 2007;176(1):4–20.
ПабМед Статья Google ученый
Thifault S, Lalonde R, Sanon N, Hamet P. Сравнения между мышами C57BL/6J и A/J по двигательной активности и координации, способности протыкать дырки и пространственному обучению. Мозг Рес Бык. 2002;58(2):213–8.
КАС пабмед Статья Google ученый
Рада-Иглесиас А., Баджпай Р., Свигут Т., Бругманн С.А., Флинн Р.А., Высоцкая Дж. Уникальная сигнатура хроматина раскрывает энхансеры раннего развития человека. Природа. 2011;470(7333):279–83.
КАС пабмед Статья Google ученый
Siersbæk R, Madsen JGS, Javierre BM, Nielsen R, Bagge EK, Cairns J, et al. Динамическая перестройка петель хроматина, закрепленных на промоторе, во время дифференцировки адипоцитов. Мол Ячейка. 2017;66(3):420-435.e5.
ПабМед Статья КАС Google ученый
Сатия Р., Фаррелл Дж. А., Геннерт Д., Шир А. Ф., Регев А. Пространственная реконструкция данных экспрессии генов одноклеточных. Нац биотехнолог. 2015;33(5):495–502.
КАС пабмед ПабМед Центральный Статья Google ученый
Bennett ML, Bennett FC, Liddelow SA, Ajami B, Zamanian JL, Fernhoff NB, et al. Новые инструменты для изучения микроглии в ЦНС мыши и человека. Proc Natl Acad Sci USA. 2016;113(12):E1738–46.
КАС пабмед ПабМед Центральный Статья Google ученый
Cahoy JD, Emery B, Kaushal A, Foo LC, Zamanian JL, Christopherson KS, et al.
База данных транскриптомов астроцитов, нейронов и олигодендроцитов: новый ресурс для понимания развития и функционирования мозга. Дж. Нейроски. 2008;28(1):264–78.КАС пабмед ПабМед Центральный Статья Google ученый
Lu QR, Sun T, Zhu Z, Ma N, Garcia M, Stiles CD, et al. Общая потребность развития в функции olig указывает на связь двигательных нейронов / олигодендроцитов. Клетка. 2002;109(1):75–86.
КАС пабмед Статья Google ученый
Мэй Ф., Ван Х., Лю С., Ню Дж., Ван Л., Хе Ю. и др. Специфическая для стадии делеция Olig2 передает противоположные функции дифференцировки и созревания олигодендроцитов. Дж. Нейроски. 2013;33(19): 8454–62.
КАС пабмед ПабМед Центральный Статья Google ученый
McLean CY, Bristor D, Hiller M, Clarke SL, Schaar BT, Lowe CB, et al.
GREAT улучшает функциональную интерпретацию цис-регуляторных регионов. Нац биотехнолог. 2010;28(5):495–501.КАС пабмед ПабМед Центральный Статья Google ученый
Ginhoux F, Greter M, Leboeuf M, Nandi S, See P, Gokhan S, et al. Анализ картирования судеб показывает, что взрослая микроглия происходит от примитивных макрофагов. Наука. 2010;330(6005):841–5.
КАС пабмед ПабМед Центральный Статья Google ученый
Stolt CC, Lommes P, Sock E, Chaboissier MC, Schedl A, Wegner M. Фактор транскрипции Sox9 определяет выбор судьбы глии в развивающемся спинном мозге. Гены Дев. 2003;17(13):1677–89.
КАС пабмед ПабМед Центральный Статья Google ученый
Госселин Д., Скола Д., Куфаль Н.Г., Холтман И.Р., Шлахецки Дж.
К.М., Сайти Э. и др. Зависимая от окружающей среды транскрипционная сеть определяет идентичность микроглии человека. Наука. 2017. https://science.sciencemag.org/content/early/2017/05/24/science.aal3222. По состоянию на 22 мая 2020 г.Кирдорф К., Эрни Д., Гольдманн Т., Сандер В., Шульц С., Пердигуэро Э.Г. и другие. Микроглия возникает из эритромиелоидных предшественников через Pu.1- и Irf8-зависимые пути. Нат Нейроски. 2013;16(3):273–80.
КАС пабмед Статья Google ученый
Лю М., Чжан Л., Марсбум Г., Джамбусариа А., Сюн С., Тот П.Т. и др. Sox17 необходим для регенерации эндотелия после воспалительного повреждения сосудов. Нац коммун. 2019;10(1):2126.
ПабМед ПабМед Центральный Статья КАС Google ученый
Stolt CC, Rehberg S, Ader M, Lommes P, Riethmacher D, Schachner M, et al.
Терминальная дифференцировка миелинобразующих олигодендроцитов зависит от транскрипционного фактора Sox10. Гены Дев. 2002;16(2):165–70.КАС пабмед ПабМед Центральный Статья Google ученый
Денин Б., Хо Р., Лукашевич А., Хохстим С.Дж., Гроностайски Р.М., Андерсон Д.Дж. Фактор транскрипции NFIA контролирует начало глиогенеза в развивающемся спинном мозге. Нейрон. 2006;52(6):953–68.
КАС пабмед Статья Google ученый
Wilczynska KM, Singh SK, Adams B, Bryan L, Rao RR, Valerie K, et al. Изоформы ядерного фактора I регулируют экспрессию генов во время дифференцировки нейронных предшественников человека в астроциты. Стволовые клетки. 2009 г.;27(5):1173–81.
КАС пабмед ПабМед Центральный Статья Google ученый
Маткович-Натан О.
, Винтер Д.Р., Гилади А., Агилар С.В., Спинрад А., Саррацин С. и др. Развитие микроглии следует поэтапной программе регулирования гомеостаза мозга. Наука. 2016;353(6301). https://science.sciencemag.org/content/353/6301/aad8670. По состоянию на 22 мая 2020 г.Sugiaman-Trapman D, Vitezic M, Jouhilahti EM, Mathelier A, Lauter G, Misra S, et al. Характеристика семейства факторов транскрипции RFX человека с помощью анализа регуляторных и целевых генов. Геномика BMC. 2018;19(1):181.
ПабМед ПабМед Центральный Статья КАС Google ученый
Пирс С.Э., Тайсон Т., Бумс А., Прал Дж., Кутзи Г.А. Генетический риск болезни Паркинсона в линии нейронов среднего мозга. Нейробиол Дис. 2018;114:53–64.
КАС пабмед Статья Google ученый
Allhoff M, Seré K, Pires JF, Zenke M, Costa IG.
Дифференциальный пиковый вызов сигналов ChIP-seq с повторами с THOR. Нуклеиновые Кислоты Res. 2016;44(20):e153.ПабМед ПабМед Центральный Google ученый
Моралес М., Марголис Э.Б. Вентральная область покрышки: клеточная неоднородность, связность и поведение. Нат Рев Нейроски. 2017;18(2):73–85.
КАС Статья пабмед Google ученый
Ponder CA, Kliethermes CL, Drew MR, Muller J, Das K, Risbrough VB, et al. Отбор на контекстуальное обусловливание страха влияет на тревожное поведение и экспрессию генов. Гены Мозг Поведение. 2007;6(8):736–49..
КАС пабмед Статья Google ученый
Португалия Г.С., Уилкинсон Д.С., Кенни Дж.В., Салливан С., Гулд Т.Дж. Зависимые от напряжения эффекты острого, хронического никотина и отмены хронического никотина на обусловленность страха.
Поведение Жене. 2012;42(1):133–50.ПабМед Статья Google ученый
Йонеяма Н., Крэбб Дж. К., Форд М. М., Мурильо А., Финн Д. А. Добровольное потребление этанола у 22 инбредных линий мышей. Алкоголь. 2008;42(3):149–60.
КАС пабмед ПабМед Центральный Статья Google ученый
Nusse R, Clevers H. Передача сигналов Wnt/β-катенина, болезнь и новые терапевтические методы. Клетка. 2017;169(6):985–99.
КАС пабмед Статья Google ученый
Мао CD, Byers SW. Зависимая от клеточного контекста экспрессия и функция TCF/LEF: альтернативные рассказы о потенциалах репрессии, дерепрессии и активации. Crit Rev Eukaryot Gene Expr. 2011;21(3):207–36.
КАС пабмед ПабМед Центральный Статья Google ученый
Wu TP, Wang T, Seetin MG, Lai Y, Zhu S, Lin K, et al. Метилирование ДНК N 6-аденина в эмбриональных стволовых клетках млекопитающих. Природа. 2016;532(7599): 329–33.
КАС пабмед ПабМед Центральный Статья Google ученый
Arnold CD, Gerlach D, Stelzer C, Boryń LM, Rath M, Stark A. Полногеномные количественные карты энхансерной активности, идентифицированные с помощью STARR-seq. Наука. 2013;339(6123):1074–1077.
КАС пабмед Статья Google ученый
McKay DJ, Lieb JD. Общий набор регуляторных элементов ДНК формирует придатки дрозофилы.
Ячейка Дев. 2013;27(3):306–18.КАС пабмед Статья Google ученый
Лара-Астиасо Д., Вайнер А., Лоренцо-Вивас Э., Зарецкий И., Джайтин Д.А., Дэвид Э. и др. Динамика состояния хроматина при кроветворении. Наука. 2014;345(6199):943–9.
КАС пабмед ПабМед Центральный Статья Google ученый
Палфи М., Шульце Г., Вален Э., Вастенхаув Н.Л. Доступность хроматина установлена Pou5f3, Sox19b и Nanog запускает гены для активации во время активации генома рыбок данио. Генетика PLoS. 2020;16(1):e1008546.
ПабМед ПабМед Центральный Статья КАС Google ученый
McGary KL, Park TJ, Woods JO, Cha HJ, Wallingford JB, Marcotte EM. Систематическое открытие неочевидных моделей болезней человека с помощью ортологических фенотипов.
Proc Natl Acad Sci. 2010;107(14):6544–9.КАС пабмед ПабМед Центральный Статья Google ученый
Lodato MA, Ng CW, Wamstad JA, Cheng AW, Thai KK, Fraenkel E, et al. SOX2 совместно занимает дистальные энхансерные элементы с различными факторами POU в ESCs и NPC, чтобы специфицировать состояние клетки. ПЛОС Жене. 2013;9(2):e1003288.
КАС пабмед ПабМед Центральный Статья Google ученый
Фэй Дж. К., Маккалоу Х.Л., Снеговски П.Д., Эйзен М.Б. Популяционная генетическая изменчивость в экспрессии генов связана с фенотипической изменчивостью в Saccharomyces cerevisiae . Геном биол. 2004;5(4):R26.
ПабМед ПабМед Центральный Статья Google ученый
Keane TM, Goodstadt L, Danecek P, White MA, Wong K, Yalcin B, et al.
Геномная изменчивость мыши и ее влияние на фенотипы и регуляцию генов. Природа. 2011;477(7364):289–94.КАС пабмед ПабМед Центральный Статья Google ученый
Massouras A, Waszak SM, Albarca-Aguilera M, Hens K, Holcombe W, Ayroles JF, et al. Геномная изменчивость и ее влияние на экспрессию генов у Drosophila melanogaster. ПЛОС Жене. 2012;8(11):e1003055.
КАС пабмед ПабМед Центральный Статья Google ученый
Стори Дж.Д., Мэдой Дж., Страут Дж.Л., Вурфель М., Рональд Дж., Эйки Дж.М. Изменчивость экспрессии генов внутри и между человеческими популяциями. Am J Hum Genet. 2007; 80 (3): 502–9..
КАС пабмед ПабМед Центральный Статья Google ученый
Цао Дж., Кусанович Д.А., Рамани В., Агамирзайе Д., Плинер Х.
А., Хилл А.Дж. и др. Совместное профилирование доступности хроматина и экспрессии генов в тысячах отдельных клеток. Наука. 2018;361(6409):1380–5.КАС пабмед ПабМед Центральный Статья Google ученый
Ge D, Fellay J, Thompson AJ, Simon JS, Shianna KV, Urban TJ, et al. Генетическая изменчивость IL28B предсказывает элиминацию вируса, вызванную лечением гепатита С. Природа. 2009 г.;461(7262):399–401.
КАС пабмед Статья Google ученый
Grundberg E, Small KS, Hedman ÅK, Nica AC, Buil A, Keildson S, et al. Картирование цис- и трансрегуляторных эффектов во многих тканях у близнецов. Нат Жене. 2012;44(10):1084–9.
КАС пабмед ПабМед Центральный Статья Google ученый
Онмахт Дж., Мэй П., Синкконен Л., Крюгер Р.
Отсутствие наследственности при болезни Паркинсона: новая роль некодирующих генетических вариаций. J Neural Transm. 2020 г. https://doi.org/10.1007/s00702-020-02184-0.Артикул пабмед Google ученый
Кроули Дж.Дж., Жаботинский В., Сунь В., Хуан С., Пакатчи И.К., Ким Й. и др. Анализ экспрессии аллель-специфического гена в сильно дивергентных скрещиваниях мышей выявляет распространенный аллельный дисбаланс. Нат Жене. 2015;47(4):353–60.
КАС пабмед ПабМед Центральный Статья Google ученый
Nott A, Holtman IR, Coufal NG, Schlachetzki JCM, Yu M, Hu R, et al. Карты интерактома энхансер-промотор, специфичные для типа клеток мозга, и ассоциация риска заболевания. Наука. 2019;366(6469):1134–9.
КАС пабмед ПабМед Центральный Статья Google ученый
Albert FW, Bloom JS, Siegel J, Day L, Kruglyak L. Генетика трансрегуляторных вариаций экспрессии генов. Wittkopp PJ, редактор. электронная жизнь. 2018;7:e35471.
Лю Х, Ли Йи, Притчард Дж.К. Транс-воздействие на экспрессию генов может привести к омнигенному наследованию. Клетка. 2019;177(4):1022-1034.e6.
КАС пабмед ПабМед Центральный Статья Google ученый
Бем Дж., Брожко Н., Чакраборти С., Липец М.А., Козинский К., Нагальский А. и другие. Передача сигналов Wnt/β-катенина при развитии мозга и психических расстройствах: не забывая о TCF7L2.
ФЭБС лат. 2019;593(13):1654–74.КАС пабмед ПабМед Центральный Статья Google ученый
Winham SJ, Cuellar-Barboza AB, Oliveros A, McElroy SL, Crow S, Colby C, et al. Полногеномное ассоциативное исследование биполярного расстройства с учетом влияния индекса массы тела выявило новый аллель риска в TCF7L2. Мол Психиатрия. 2014;19(9):1010–6.
КАС пабмед Статья Google ученый
Sittig LJ, Carbonetto P, Engel KA, Krauss KS, Barrios-Camacho CM, Palmer AA. Генетический фон ограничивает обобщаемость отношений генотип-фенотип. Нейрон. 2016;91(6):1253–9.
КАС пабмед ПабМед Центральный Статья Google ученый
Corces MR, Trevino AE, Hamilton EG, Greenside PG, Sinnott-Armstrong NA, Vesuna S, et al. Улучшенный протокол ATAC-seq уменьшает фон и позволяет исследовать замороженные ткани.
Нат Методы. 2017;14(10):959–62.КАС пабмед ПабМед Центральный Статья Google ученый
Ли З., Шульц М.Х., Лук Т., Бегеманн М., Зенке М., Коста И.Г. Идентификация сайтов связывания факторов транскрипции с использованием ATAC-seq. Геном биол. 2019;20(1):45.
ПабМед ПабМед Центральный Статья Google ученый
Фэн Дж., Лю Т., Цинь Б., Чжан Ю., Лю Х.С. Идентификация обогащения ChIP-seq с использованием MACS. Нат Проток. 2012;7(9): 1728–40.
КАС пабмед Статья Google ученый
Хайнц С., Беннер С., Спанн Н., Бертолино Э., Лин Ю.С., Ласло П. и др. Простые комбинации транскрипционных факторов, определяющих происхождение, задают цис-регуляторные элементы, необходимые для идентичности макрофагов и В-клеток. Мол Ячейка.
2010;38(4):576–89.КАС пабмед ПабМед Центральный Статья Google ученый
Эндрюс С. FastQC: инструмент контроля качества для высокопроизводительных данных последовательностей. 2010.
Schubert M, Ermini L, Sarkissian CD, Jónsson H, Ginolhac A, Schaefer R, et al. Характеристика древних и современных геномов путем обнаружения SNP и филогеномного и метагеномного анализа с использованием PALEOMIX. Нат Проток. 2014;9(5):1056–82.
КАС пабмед Статья Google ученый
Li H, Handsaker B, Wysoker A, Fennell T, Ruan J, Homer N, et al. Формат выравнивания/карты последовательностей и SAMtools. Биоинформатика. 2009 г.;25(16):2078-9.
ПабМед ПабМед Центральный Статья КАС Google ученый
Короткова ТМ, Пономаренко АА, Браун РЭ, Хаас ХЛ. Функциональное разнообразие дофаминовых и ГАМКергических нейронов вентрального среднего мозга. Мол Нейробиол. 2004;29(3):243–59.
КАС пабмед Статья Google ученый
Gui Y, Thomas MH, Garcia P, Karout M, Halder R, Michelucci A, et al. Ген 1, трансформирующий опухоль гипофиза, управляет регуляторными вариациями генов в вентральной части среднего мозга мышей во время старения. Фронт Жене. 2020 г. https://doi.org/10.3389/fgene.2020.566734.
Артикул пабмед ПабМед Центральный Google ученый
Батлер А., Хоффман П., Смиберт П., Папалекси Э., Сатия Р. Интеграция транскриптомных данных отдельных клеток в различных условиях, технологиях и видах. Нац биотехнолог. 2018. https://www.nature.com/articles/nbt.4096. По состоянию на 3 апреля 2018 г.
Liu K, Pan C, Kuhn A, Nievergelt AP, Fantner GE, Milenkovic O, et al. Обнаружение топологических вариаций ДНК на уровне одной молекулы. Нац коммун. 2019;10(1):3.
КАС пабмед ПабМед Центральный Статья Google ученый
Bogue MA, Philip VM, Walton DO, Grubb SC, Dunn MH, Kolishovski G, et al. База данных фенотипов мышей: репозиторий данных и набор для анализа тщательно подобранных данных о первичных фенотипах мышей. Нуклеиновые Кислоты Res. 2020;48(Д1):Д716–23.
КАС пабмед Google ученый
Адамс М.Д., Целникер С.Э., Холт Р.А., Эванс К.А., Гокейн Д.Д., Аманатиды П.Г. и др. Последовательность генома Drosophila melanogaster. Наука. 2000;287(5461):2185–95.
ПабМед Статья Google ученый
Скачать ссылки
Благодарности
Мы хотели бы поблагодарить Drs. Орельен Жинолхак и Антула Геньо за поддержку биоинформатического анализа, д-р Джалил Кувар (Ветеринарный центр Люксембургского университета) за помощь в разведении экспериментальных мышей, д-р Раши Гальдер из центра секвенирования LCSB за высокопроизводительное секвенирование и Серхио Хельгета за Помогите с выделением ядер. Вычислительный анализ, представленный в этой статье, был выполнен с использованием средств HPC Люксембургского университета.
Финансирование
LS и MB выражают благодарность Люксембургскому национальному исследовательскому фонду (FNR) за поддержку (грант FNR CORE C15/BM/10406131). LS и JO хотели бы поблагодарить Fondation du Pélican de Mie et Pierre Hippert-Faber и Luxembourg Rotary Foundation за финансирование. MHT, PG и MB выражают благодарность профессору М. Миттельбронну, финансируемому грантом FNR PEARL P16/BM/111, за поддержку.
Информация об авторе
Авторы и организации
Факультет наук о жизни и медицины (DLSM), Люксембургский университет, Бельво, Люксембург
Yujuan Gui, Jochen Ohnmacht, Thomas Sauter & Lasse Sinkkonen
Люксембургский центр системной биомедицины (LCSB), Люксембургский университет, Бельво, Люксембург
Kamil Grzyb, Ohnma Puel, Jélanie Garchen Buttini & Alexander Skupin
Авторы
- Yujuan Gui
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Академия
- Kamil Grzyb
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
- Мелани Х. Томас
Просмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
- Jochen Ohnmacht
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Pierre Garcia
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Manuel Buttini
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Александр Скупин
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
- Томас Заутер
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
- Lasse Sinkkonen
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
Contributions
YG, MB и LS задумали проект. YG, KG, JO, AS, TS и LS разработали эксперименты. YG и KG выполнили snATAC-seq. YG и JO выполнили объемную ATAC-seq, а YG провела эксперименты ChIP-seq. Мышиные ткани, приготовленные MHT и PG. YG провел весь биоинформационный анализ с помощью TS и LS. YG и LS проанализировали результаты. MB, TS и LS предоставили финансирование. YG и LS написали рукопись. Все авторы прочитали и одобрили окончательный вариант рукописи.
Автор, ответственный за переписку
Переписка с Лассе Синкконен.
Декларация этики
Утверждение этики и согласие на участие
Протокол животных был одобрен Комитетом по этике экспериментов на животных (AEEC) в соответствии с национальными рекомендациями закона о защите животных в Люксембурге ( Регламент великого герцога , принятый в январе 11 января 2013 г.).
Согласие на публикацию
Не применимо.
Конкурирующие интересы
Авторы заявляют об отсутствии конкурирующих интересов.
Дополнительная информация
Примечание издателя
Springer Nature остается нейтральной в отношении юрисдикционных претензий в опубликованных картах и институциональной принадлежности.
Дополнительная информация
Дополнительный файл 1: Рисунок S1.snATAC-seq на вентральной части среднего мозга C57BL/6J и A/J выявил доступность хроматина, специфичного для типа клеток. Рисунок S2. h4K27ac ChIP-seq и ATAC-seq, коррелирующие с экспрессией генов. Рисунок S3. Дифференциальные пики могут выявить специфические для штамма ТФ.
Дополнительный файл 2: Таблица S1.Композиция клеточного типа в вентральной части среднего мозга C57BL/6J и A/J в snATACseq. Типы клеток и идентифицированные номера клеток указаны для обоих штаммов.
Дополнительный файл 3 : Таблица S2. Гены идентичности клеточного типа, определенные из существующей scRNA-seq. Официальные символы генов для идентифицированных генов клеточной идентичности перечислены под каждым соответствующим типом клеток.
: Таблица S3. Анализ обогащения генов идентичности клеточного типа, определенных из существующей scRNA-seq. Представлены результаты обогащения GO для генов клеточной идентичности для каждого типа клеток. Анализ обогащения был выполнен с использованием идентификаторов терминов Enrichr и GO, значения обогащения p , баллы и идентифицированные имена генов предоставлены для каждого термина GO.
Дополнительный файл 5: Таблица S4. Пики идентичности клеточного типа, определенные путем связывания специфических для клеточного типа пиков с регуляторными областями генов идентичности клеточного типа. Координаты хромосом для всех пиков идентичности клеток предоставляются для каждого типа клеток.
Дополнительный файл 6 : Таблица S5. Предполагаемые регуляторные варианты. Координаты хромосом и основные/альтернативные аллели приведены для 3909 вариантов, расположенных в пределах следов TF в активных энхансерах и обозначенных как предполагаемые регуляторные варианты.
: Таблица S6. Дифференциально экспрессируемые гены из последовательностей последовательностей РНК вентральной части среднего мозга между C57BL/6J и A/J. Каждый штамм имеет 12 повторностей (6 самцов и 6 самок). Дифференциальные гены определяются как 92 116 p 92 117 adj < 0,05.
Дополнительный файл 8: Таблица S7. Дифференциальные пики h4K27ac между C57BL/6J и A/J. Хромосомные координаты пиков и p — для каждого дифференциального пика указаны значения знаковой разницы между штаммами ( p < 1 x 10 -18 ).
Дополнительный файл 9 : Таблица S8. Предполагаемые вышестоящие регуляторы между C57BL/6J и A/J. Предсказанные вышестоящие регуляторы получены из IPA на основе дифференциально экспрессируемых генов из объемной РНК-последовательности среднего мозга между C57BL/6J и A/J ( p adj <0,05, log2FC>1).
Права и разрешения
Открытый доступ Эта статья находится под лицензией Creative Commons Attribution 4.0 International License, которая разрешает использование, совместное использование, адаптацию, распространение и воспроизведение на любом носителе или в любом формате, при условии, что вы укажете соответствующую ссылку на оригинальный автор(ы) и источник, предоставьте ссылку на лицензию Creative Commons и укажите, были ли внесены изменения. Изображения или другие сторонние материалы в этой статье включены в лицензию Creative Commons на статью, если иное не указано в кредитной строке материала. Если материал не включен в лицензию Creative Commons статьи, а ваше предполагаемое использование не разрешено законом или выходит за рамки разрешенного использования, вам необходимо получить разрешение непосредственно от правообладателя. Чтобы просмотреть копию этой лицензии, посетите http://creativecommons.org/licenses/by/4.0/. Отказ Creative Commons от права на общественное достояние (http://creativecommons.
